
Neuroscience Research Booklet
Neuroscience Research Related Cytokine Products
Introduction
Neuroscience is the biological study of the nervous system, encompassing those scientific disciplines concerned with the development, molecular and cellular structure, chemistry, functionality, evolution, and pathology of the neural networks constructing the nervous system. The vertebrate nervous system is a sophisticated networking of neural cells that function through the transmission of excitatory or inhibitory signaling to process information and orchestrate all bodily functions, including the capacity for motor and sensory function, cognition, and emotion. Arising from the ectoderm, the most exterior of the three germ cell layers, this system is comprised of both the central and peripheral nervous systems. The central nervous system, consisting of the brain and spinal cord, functions to receive, interpret, and respond to the nerve impulses of the peripheral nervous system. The peripheral nervous system serves as the connection between the central nervous system and the body, transporting sensory and motor information to and from the central nervous system, respectively.
Constructed from those sensory and motor neurons found outside the white and gray matter of the spinal cord and brain, the peripheral nervous system is generally divided into the sensory-somatic and autonomic nervous systems; the last of which is then subdivided into the sympathetic and parasympathetic nervous system. Altogether, the nervous system is biologically comprised of the neurons and supporting cells required to control the voluntary and involuntary actions of the body, including Schwann cells, satellite cells (or ganglionic gliocytes), oligodendrocytes, microglia, astrocytes, and ependymal cells. Constituting and supporting the brain and spinal cord within the central nervous system, these cells also serve to establish and support the cranial and spinal nerves of the peripheral nervous system, which arise from the brain and spinal cord, respectively. Understanding the development and homeostasis of the nervous system, the many pathways responsible for maintaining and regulating its proper functionality, and the implications of nervous system dysfunction, remain poignant within research concerning the nervous system and neurodegenerative diseases.
Research exploring the biological processes of the nervous system and their influences on both human behavior and function offers vast possibilities for the understanding, as well as treatment, of various neurological diseases and disorders, such as Alzheimer's disease, Multiple Sclerosis, Parkinson's disease and Epilepsy.
This booklet describes a diverse number of commercially available cytokine products related to neuroscience research. By offering this wide array of cytokine reagents, it is PeproTech's goal to contribute to the continuing advances in neuroscience related research and the overall improvement of worldwide health.
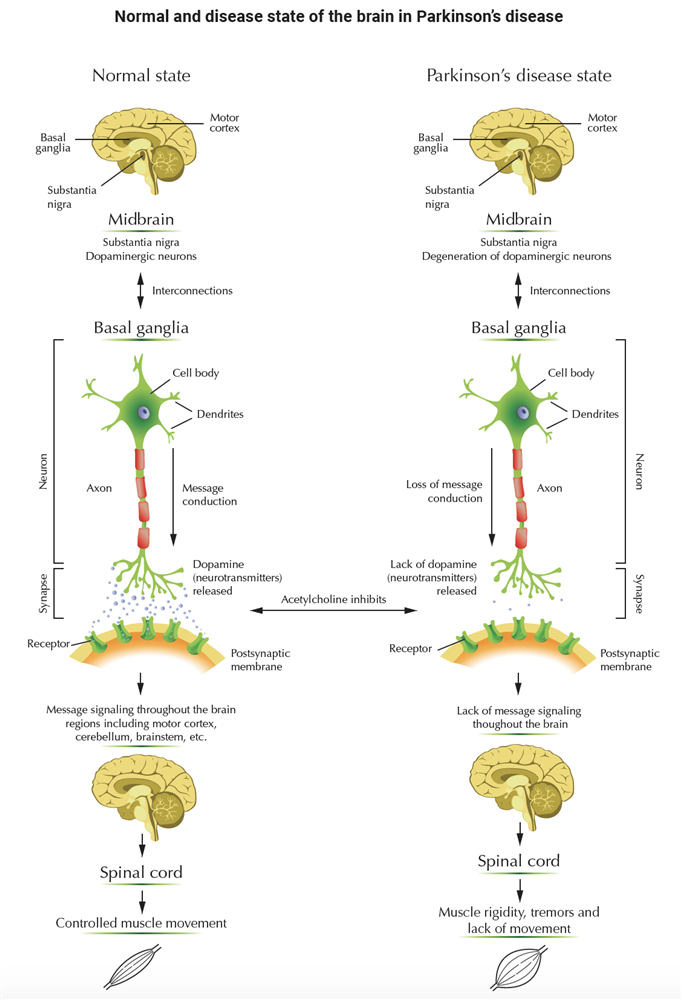
Alzheimer’s Disease and Inflammation
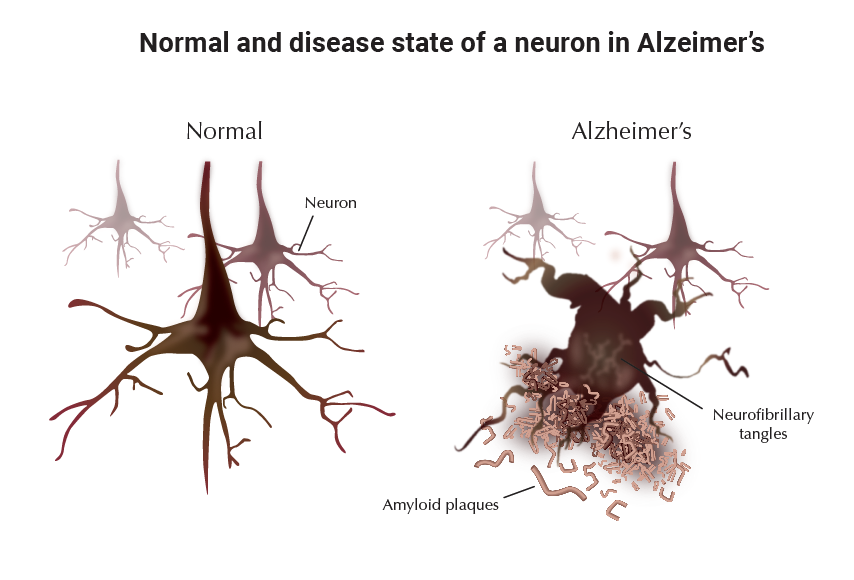
Alzheimer's disease (AD) is a progressive neurodegenerative disorder marked by accumulation of extracellular deposits of beta-amyloid in brain regions that are important for memory and cognition (e.g. the hippocampus and cerebral cortex). Beta-amyloid (Aβ) is comprised of 40 and 42 amino-acid peptides (Aβ40 and Aβ42), generated by proteolytic processing of a widely expressed cell surface protein called amyloid precursor protein (APP). Aβ, in particular Aβ42, is prone to concentration-dependent oligomerization and aggregation. Rising levels of Aβ in the cerebrospinal fluid gradually lead to formation of small oligomers followed by growth into protofibrils and fibrils, which can associate with other peptides and proteins to form highly insoluble neuritic plaques (also called senile plaques). The buildup of Aβ aggregates in the AD brain is followed by formation of intracellular neurofibrillary tangles and activation of local inflammatory reactions. These brain changes ultimately lead to a widespread loss of synapses, neuronal degeneration, and neurotransmitter deficits.
The sporadic form of AD usually begins after the age of 60 years, and is the most common cause of dementia in the elderly. Its prevalence rises steadily with age, affecting less than 3% of persons between ages 60 and 70, up to 12% of those between 70 and 80, and more than 40% of those over 85. Besides age, the greatest risk factor for AD is inheritance of one or two ε4 alleles of the single human apolipoprotein E gene (chromosome 19). The ε4 genetic variation increases the likelihood of developing AD 3-8 fold and lowers the age of onset by 5-10 years, as compared with ε3 homozygotes. The familial form of AD, which accounts for less than 10% of AD cases, typically begins during the fifth decade of life. It is caused by inherited mutations in the genes for APP (chromosome 21), presenilin (PS)-1 (chromosome 14), and PS-2 (chromosome 1). Warning signs of AD are subtle and include mild forgetfulness and difficulty in identifying familiar smells. The actual development of AD is marked by progressive decline in memory and language function, personality changes, and finally dementia. Death usually occurs as a result of minor respiratory complications in the middle of the night. In the year 2000, there were an estimated 4.5 million people in the U.S. with AD, and this number is predicted to triple by 2050, if no therapy intervenes.
APP and Generation of AβPlaques
Amyloid precursor protein (APP) occurs in 3 major isoforms containing 770, 751, or 695 amino-acid residues, arising from alternative splicing of a single gene. The main difference between these isoforms is the presence or absence of a 51-residue Kunitz protease inhibitor (KPI) domain. Interestingly, APP-695 which lacks the KPI domain is expressed exclusively in neurons and at higher levels than the KPI-containing forms (APP-751 and APP-770). The latter are widely expressed in non-neuronal cells throughout the body, including astrocytes, microglia, and other brain cells. The APP gene is highly conserved in evolution, and nearly identical in mammalian species. It encodes a single transmembrane polypeptide whose primary structure contains a 17-residue signal peptide for secretion, a large extracellular N-terminal domain, a 24-residue transmembrane domain, and a cytoplasmic C-terminal tail of 47 amino-acid residues (Figure 1). The Aβ sequence, marked in red in figure 1, lies partially outside the cell membrane (amino acids 1-28 of Aβ) and partially within the membrane (amino acids 29-40 and 29-42 of Aβ40 and Aβ42, respectively). During normal metabolism, APP, whose half-life is less than an hour, can be processed by three proteolytic activities, designated α−, β−, and γ-secretase (Table I).
Figure 1: Proteolytic Processing of APP
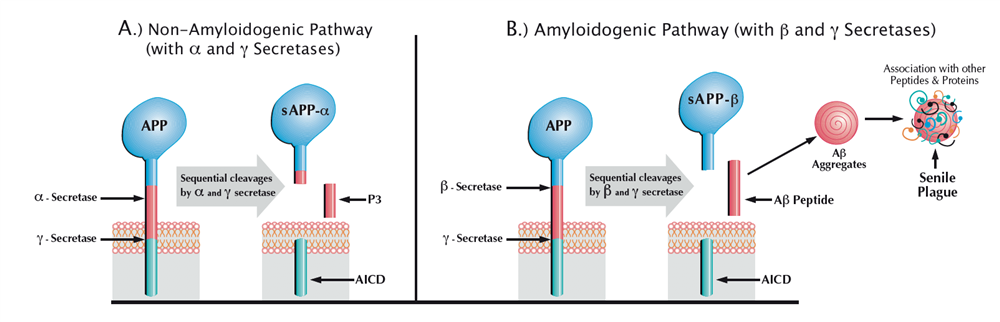
Table 1: Proteolytic Processing of APP
| α-secretase has not been identified as any single proteinase, but three members of the ADAM (α disintegrin and metalloproteinase) family, ADAM-9, ADAM-10, and ADAM-17 (TACE) are candidate α-secretases. Up-regulation of α-secretase activity reduces Aβ formation, and is being tested as a possible therapeutic treatment for AD. |
| β-secretase also known as BACE1 (β-site APP cleaving enzyme) is a unique member of the pepsin family of aspartyl proteinases. It is a type I transmembrane protein whose absence in BACE knockout mice has no adverse consequences, but dramatically reduces levels of Aβ. Currently, BACE1 is a major target for development of inhibitors to treat AD. |
| γ-secretase is a membrane protein complex comprised of presenilin homodimer (PS1 or PS2), nicastrin, Aph1 (Aph1a or Aph1b) and Pen2. Such minimal complex is sufficient for γ-secretase activity, although other components may exist. γ-Secretase is predominantly located in the endoplasmic reticulum and the cis-Golgi, and is capable of cleaving intra-membrane peptide bonds in a variety of additional proteins, including Notch, E-cadherin, and ErbB4. |
Cleavage of APP by α-secretase, which splits the peptide bond between residues 16 and 17 of the Aβ sequence, precludes Aβ formation and is referred to as the non-amyloidogenic pathway. It leads to release of a large soluble extracellular fragment (sAPP-α) and retention of an 83-residue membrane-bound fragment (C83). Alternatively, APP can be cleaved by β-secretase, which cuts at the N-terminus of Aβ, thus generating a slightly smaller extracellular fragment (sAPP-β) and a 99-residue membrane-bound fragment (C99). Subsequent to their formation, C83 and C99 undergo conformational change and become targets for the presenilin (PS1/PS2)-dependent γ-secretase, which cleaves them within their plasma membrane domain. This unusual proteolysis is heterogeneous and yields peptides with slightly different lengths. It generates 24 and 26 amino-acid peptides (P3) from C83, and Aβ40 and Aβ42 from C99. The cleavage of either C83 or C99 by γ-secretase also yields an amyloid intracellular domain (AICD) of 57-59 amino-acid residues, which moves to the nucleus where it may modulate transcription of target genes. Although neither the physiological role of APP nor that of its proteolytically-derived fragments has been demonstrated directly, several lines of evidence suggest that at least one of the γ-secretase-derived fragments may play an important neuroprotective role. For example, mice whose cerebral γ-secretase activity was eliminated postnatally by combined genetic ablation of both PS1 and PS2 genes developed age-dependent cognitive deficits and neurodegeneration. As expected, amyloid plaques, the pathological hallmark of AD, were absent in the brain of these animals (1).
The steady-state levels of Aβ in the cerebrospinal fluid and plasma of healthy individuals are 3-8 nM and less than 0.5 nM, respectively. Normally, the Aβ pool contains predominantly Aβ40, which is much less prone to aggregation than Aβ42. In familial AD, the Aβ42/Aβ40 ratio is considerably higher than normal as a result of mutated forms of APP, PS1, and PS2, which share a common feature of modulating γ-secretase cleavage to increase production of the highly amyloidogenic Aβ42 peptide. Overexpression of normal APP due to duplication of chromosome 21 (Down's syndrome) results in increased secretion of Aβ from birth and invariably leads to premature AD, usually during the fourth decade of life. An interesting exception is a rare case of a Down's syndrome patient whose chromosome 21 was diploid for the APP gene, and who lived to the age of 78 years with no signs of dementia (2).
Aβ-induced Inflammation
Inflammation is a bodily response to irritation or injury, aimed at eliminating both foreign and endogenously-derived contaminating agents. The pivotal cells involved in this activity are blood polymorphonuclear leukocytes (PMNLs) and monocyte-derived tissue macrophages. These cells, dubbed ìthe professional phagocytesî, are capable of ingesting and degrading particulate matter. Their phagocytic activity is often accompanied by release and/or leakage of oxidants (e.g. nitric oxide, hydrogen peroxide, and hydroxyl radicals), which are implicated in a variety of tissue damage mechanisms. The recognition signal for ingestion may be either particle-surface determinants, or certain serum proteins, called opsonins, which bind to the particle and render it ingestible. Circulating PMNLs are highly specialized to engulf and kill invading microorganisms; however they often die shortly thereafter, leaving puss behind. Macrophage-mediated responses, on the other hand, are typically more durable and less harsh than PMNL responses, and can accomplish degradation and recycling of accumulated debris. Normally, the immediate and early response to tissue injury is acute and involves massive infiltration with PMNL (mainly neutrophils). The cardinal signs of this reaction are redness and swelling with heat and pain (ìrubor et tumor cum calore et doloreî). The PMNL-mediated response is usually phased out within hours, giving way to repair processes to heal and reconstitute the sites of injury. If clearance of irritants is incomplete, inflammation can persist indefinitely, primarily as a low-grade macrophage-mediated response. The collateral damage caused by this type of inflammation usually accumulates slowly, sometimes asymptomatically for years, and if unabated, can lead to severe tissue deterioration. Accelerated tissue damage results from chronic inflammation that has the potential to erupt episodically, as in Gout and Crohn's disease. In recent years it has become generally accepted that low-grade chronic inflammation is a silent killer, underlying severe human diseases including atherosclerosis, colorectal cancer, and Alzheimer's disease.
Although the precise relationship between inflammation and the pathogenesis of AD remains unclear, there is compelling evidence that neuroinflammation is critical for driving the disease process. Numerous epidemiological and retrospective studies have demonstrated that long term use of nonsteroidal anti-inflammatory drugs (NSAIDs) significantly reduces the risk for developing AD (3). During the past 15 years, a wide range of inflammatory mediators have been found to be upregulated in the AD brain, including complement components, acute phage reactants, oxidative stress enzymes, and inflammatory cytokines. The presence of these mediators in the AD brain is associated with abundance of activated microglia and astrocytes in regions with high Aβ-plaque burden. Microglial cells, which constitute 10-15% of the cellular population in the brain, have a monocytic origin and appear to possess a macrophage-like-phagocytic capacity. The accumulation of activated monocyte-derived cells around insoluble deposits of phagocytosis-resistant material is a common phenomenon in peripheral amyloidogenesis. Such a response has been defined as inflammatory over a century ago. However, unlike inflammatory responses in other tissues and organs, neuroinflammation is exclusively mediated by resident brain cells. The blood-brain barrier (BBB), which separates the central nervous system (CNS) from the blood, prevents not only harmful agents such as microbes and viruses but also circulating leukocytes and antibodies from entering into the brain. Consequently, the CNS can sustain only low-grade chronic inflammation, which is primarily mediated by activated microglia. Resting microglial cells play an important neurotrophic role, regulate various metabolic processes, and help to maintain homeostasis in the CNS. Of particular relevance to the pathogenesis of AD is the role of microglial-derived apoE, which is commonly associated with Aβ deposits. ApoE is a 34 kDa protein of 299 amino-acid residues, occurring as three major isoforms that differ at two residues: apoE2 (Cys112, Cys158), apoE3 (Cys112, Arg158), and apoE4 (Arg112, Arg158). ApoE is a key component of most lipoproteins (e.g. HDL and LDL), and plays an essential role in the redistribution of cholesterol and other lipids throughout the body. In the brain, apoE appears to inhibit Aβ-induced neurotoxicity and inflammation, and to promote clearance of amyloid deposits (4−7). ApoE is capable of solubilizing hydrophobic compounds by incorporating them into its core lipid-binding domain and delivering them to target cells via apoE receptors. However, apoE, especially the E4 isoform, is susceptible to proteolytic cleavage by a chymotrypsin-like protease, which removes a 27-residue C-terminal domain and generates a truncated apoE protein (apoE-272) with an exposed lipid-binding domain (8). When encountering hydrophobic Aβ assemblies, ApoE-272 is not only unable to solubilize them, but sticks to them and enhances their aggregation into highly insoluble plaques. Transgenic mice expressing high levels of apoE4-272 died at 2-4 months of age, and their brain cortex and hippocampus displayed AD-like alterations (8).
Aggregated forms of Aβ peptides induce transformation of resting microglia and astrocytes into activated cells, both in-vitro and in-vivo (9). These activated cells can express a variety of inflammatory mediators including chemokines, cytokines, complement and other acute-phase proteins. Interestingly, recent studies have demonstrated that under stressful conditions, neurons can produce a number of these factors including complement, cyclooxygenase-2 (COX-2), and inflammatory cytokines. In the vicinity of amyloid plaques, activated astrocytes undergo both hyperplasia and hypertrophy, and become large, star-shaped cells. These astrocytes form halos around the plaques, thus insulating them from nearby neurons and providing neuronal protection from toxic effects of Aβ deposits. In contrast to the peripheral positioning of astrocytes, activated microglia assume a more central location and deeply interdigitate the plaques. Accumulating evidence suggest that the initial attempt of these cells to phagocytose Aβ plaques involves limited activation of the complement pathway and generation of relatively non-neurotoxic opsonizing proteins such as the C1q and C3b complement components (10). Failure in this attempt appears to trigger further activation of the complement cascade, gradually leading to generation of more neurotoxic complement fragments and ultimately results in formation of the highly neurotoxic C5b-9 complex, also known as MAC (ìmembrane attack complexî). The presence of MAC and other complement activation proteins in the AD brain have been demonstrated by immunochemical staining procedures, indicating the significance of complement inflammation in the pathogenesis of AD (10). The perpetuation of neuroinflammation, which is accompanied by altered ionic homeostasis, free radical formation, oxidative damage, and neuritic dystrophy, may be the primary cause of neuronal death in AD. Once neuronal injury or death occurs, either due to Aβ neurotoxicity or as a result of Aβ-induced inflammation, the resulting debris may overtime create a self-amplifying cycle of neurodegenerative events.
Treating and Preventing AD
To date, there are no available therapeutic interventions that halt or reverse AD. As noted earlier, chronically taken NSAIDs appear to delay the onset of AD symptoms, probably by limiting neuronal damage caused by Aβ-induced inflammation. However, excessive use of NSAIDs targeting cyclooxygenase (COX) can cause gastrointestinal, liver, and renal toxicity. Certain NSAIDs including ibuprofen can confer protection against AD by modulating γ-secretase cleavage to lower Aβ42 production, a mechanism that does not require inhibition of COX activity (11). Derivatives of NSAIDS, which lack COX inhibitory activity but can still lower Aβ42 production, are presently being tested in patients with AD. There is evidence that maintaining low LDL levels by various cholesterol-lowering regiments decreases the risk for developing cardiovascular disease and AD. Taken together, these findings suggest that agents such as statins, which possess both cholesterol-lowering capacity and anti-inflammatory properties, could be useful anti-AD agents. The statins are FDA-approved drugs for LDL-lowering therapy, and their therapeutic usefulness in AD is currently being tested in clinical trials.
The notion that plaque formation in AD is enhanced by oxidative damage is supported by studies indicating that antioxidants can be useful inhibitors of AD. For example, high daily doses of both vitamins E and C, which possess antioxidant properties, have been reported to considerably reduce the risk of AD in elderly people (12). Another antioxidant that has been shown to possess therapeutic potential against AD is curcumin, a yellow phenolic compound found in the spice tumeric, which is a key ingredient in American mustard and Indian curry.
Figure 2: Molecular Structure of Curcumin
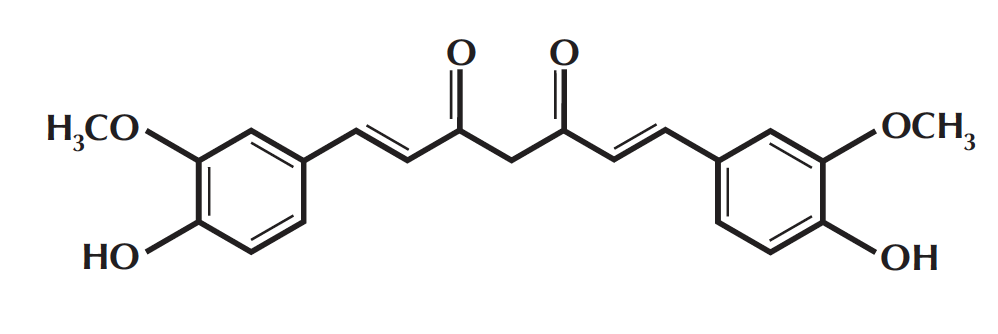
Curcumin has long been known for its potent anti-inflammatory and antioxidant activities and its widespread consumption in India is believed to be a major reason for the unusually low incidence of AD in this country, with just 1% of those 65 and older contracting the disease. In recent years, curcumin has become a subject of intense investigation because of its protective effects against human malignancies, which have been demonstrated by a number of studies. The therapeutic potential of curcumin as an anti-AD agent has recently been tested both in-vitro and in a transgenic mouse model for AD (APPsw mice) (13, 14). At concentrations of around 1 μM, curcumin was found to effectively prevent oligomerization and aggregation of Aβ peptides as well as disassemble preformed Aβ fibrils (14). When fed to aged APPsw mice with advanced amyloid levels, curcumin crossed the blood-brain barrier, bound to Aβ, and significantly reduced amyloid levels and plaque burden (13, 14). These findings suggest that the anti-inflammatory activity of curcumin stems, at least in part, from its ability to facilitate amyloid clearance. They also provide strong rationale for curcumin use in the treatment and prevention of AD, and should encourage further search for potential anti-AD agents that could be even more effective than curcumin in disaggregating Aβ fibrils.
References
(1) C.A. Saura, et al. Neuron, Vol. 42, 23-36 (2004)
(2) D.M. Walsh and D.J. Selkoe, Neuron, Vol. 44, 183-193 (2004)
(3) P.L. McGeer, et al, Neurlogy, Vol. 47, 425-432 (1996)
(4) A.M. Manelli, et al. J. Molecular Neuroscience, Vol. 23, 235-246 (2004)
(5) J. Jordan, et al. J. Neurosci., Vol. 18, 195-204 (1998)
(6) J.R. Lynch, et al. J.Neuroimmunol., Vol. 114, 107-113 (2001)
(7) D.S. Yang, et al. Neuroscience, Vol. 192, 1217-1226 (1999)
(8) F.M. Harris, et al. PNAS (USA), Vol. 100, 10966-10971 (2003)
(9) R. Strohemer and J. Rogers, J. of Alzheimer’s Disease, Vol. 3, 131-157 (2001)
(10) D.A. Loeffler, J. of Neuroinflammation, Vol. 1, 18-29 (2004)
(11) S.Weggen, et al. Nature, Vol. 414, 212-216 (2001).
(12) P.P. Zandi, et al. Arch. Neurol., Vol. 62, 82-88 (2004)
(13) G.P. Lim, et al. J. Neurosci., Vol. 21, 8370-8377 (2001)
(14) F.Yang, et al. J. Biol. Chem., Vol. 280, 5892-5901 (2005)
NEUROSCIENCE RELATED DIAGRAMS
Activin A
Activin A is a TGF-β family member that exhibits a wide range of biological activities, including regulation of cellular proliferation and differentiation, and promotion of neuronal survival. Elevated levels of Activin A in human colorectal tumors and in postmenopausal women have been implicated in colorectal and breast cancers, respectively. The biological activities of Activin A can be neutralized by inhibins and by the diffusible TGF-β antagonist, follistatin. Activin A binds to the two forms of activin receptor type I (Act RI-A and Act RI-B) and two forms of activin receptor type II (Act RII-A and Act RII-B). Activins are homodimers or heterodimers of different β subunits. They are produced as precursor proteins with an amino terminal propeptide that is cleaved to release the C-terminal bioactive ligand.
Activin B
Activin B is a TGF-β family member that exhibits a wide range of biological activities, including regulation of embryogenesis, osteogenesis, hematopoiesis, reproductive physiology and hormone secretion from the hypothalamic, pituitary and gonadal glands. Activin B, like certain other members of the TGF-β family, signals through the ActRII receptor (Activin Receptor type II). Activins are homodimers or heterodimers of different β subunits. They are produced as precursor proteins with an amino terminal propeptide that is cleaved to release the C-terminal bioactive ligand.
Adiponectin
Adiponectin is an adipose-derived secreted protein containing 236 amino acid residues. It is relatively abundant in humans and rodents, accounting for about 0.01% of total plasma protein. The circulating levels of adiponectin are decreased under conditions of obesity, insulin resistance, and type II diabetes. Disruption of adiponectin in mice causes insulin resistance and neointimal formation. Conversely, administration of recombinant adiponectin suppresses hepatic glucose production, and reverses insulin resistance associated with both lipoatrophy and obesity. The protective role of adiponectin is attributed to its anti-inflammatory properties (e.g. ability to suppress expression of TNF-α and class A scavenger receptor in macrophages).
Amphiregulin
Amphiregulin is an EGF-related growth factor that signals through the EGF/TGF-α receptor, and stimulates growth of keratinocytes, epithelial cells and some fibroblasts. Amphiregulin also inhibits the growth of certain carcinoma cell lines. Synthesized as a transmembrane protein, Amphiregulin's extracellular domain is proteolytically processed to release the mature protein. There are 6 conserved cysteine residues, which form 3 intramolecular disulfide bonds essential for biological activity.
Artemin
Artemin is a disulfide-linked homodimeric neurotrophic factor structurally related to GDNF, Artemin, Neurturin and Persephin. These proteins belong to the cysteine knot superfamily of growth factors that assume stable dimeric protein structures. Artemin, GDNF, Persephin and Neurturin all signal through a multicomponent receptor system, composed of RET (receptor tyrosine kinase) and one of the four GFRα (α1-α4) receptors. Artemin prefers the receptor GFRα3-RET, but will use other receptors as an alternative. Artemin supports the survival of all peripheral ganglia, such as sympathetic, neural crest and placodally-derived sensory neurons, and dopaminergic midbrain neurons. The functional human Artemin ligand is a disulfide-linked homodimer of two 12.0 kDa polypeptide monomers. Each monomer contains seven conserved cysteine residues, one of which is used for interchain disulfide bridging and the others are involved in intramolecular ring formation known as the cysteine knot configuration.
BDNF
BDNF is a member of the NGF family of neurotrophic growth factors. Like other members of this family, BDNF supports neuron proliferation and survival. BDNF can bind to a low affinity cell surface receptor called LNGFR, which also binds other neurotrophins such as NGF, NT-3 and NT-4. However, BDNF mediates its neurotrophic properties by signaling through a high affinity cell surface receptor called gp145/trkB. BDNF is expressed as the C-terminal portion of a 247 amino acid polypeptide precursor, which also contains a signal sequence of 18 amino acid residues and a propeptide of 110 amino acid residues.
Betacellulin
Betacellulin is an EGF-related polypeptide growth factor that signals through the EGF receptor. It is produced in several tissues, including the pancreas, small intestine, and in certain tumor cells. Betacellulin is a potent mitogen for retinal pigment epithelial cells and vascular smooth muscle cells. Human Betacellulin is initially synthesized as a glycosylated 32.0 kDa transmembrane precursor protein, which is processed by proteolytic cleavage to produce the mature sequence.
BMP-2
BMPs (Bone Morphogenetic Proteins) belong to the TGF-β superfamily of structurally related signaling proteins. BMP-2 is a potent osteoinductive cytokine, capable of inducing bone and cartilage formation in association with osteoconductive carriers such as collagen and synthetic hydroxyapatite. In addition to its osteogenic activity, BMP-2 plays an important role in cardiac morphogenesis, and is expressed in a variety of tissues, including lung, spleen, brain, liver, prostate ovary and small intestine. The functional form of BMP-2 is a 26 kDa protein composed of two identical 114 amino acid polypeptide chains linked by a single disulfide bond. Each BMP-2 monomer is expressed as the C-terminal part of a precursor polypeptide, which also contains a 23 amino acid signal sequence for secretion, and a 259 amino acid propeptide. After dimerization of this precursor, the covalent bonds between the propeptide (which is also a disulfide-linked homodimer) and the mature BMP-2 ligand are cleaved by a furin-type protease.
BMP-3
TGF-β family members are key modulators of cell proliferation, differentiation, matrix synthesis, and apoptosis. As implied by their name, BMPs initiate, promote, and regulate the development, growth, and remodeling of bone and cartilage. In addition to this role, BMPs are also involved in prenatal development and postnatal growth, remodeling and maintenance of a variety of other tissues and organs. BMP-3 is abundantly found in adult bone and, to a lesser extent, fetal cartilage. BMP-3 inhibits osteogenesis and bone formation by activating a signaling cascade that antagonizes the signaling of pro-osteogenic BMPs.
BMP-4
Bone morphogenetic proteins (BMPs) constitute a subfamily within the TGF-β superfamily of structurally related signaling proteins. Members of this superfamily are widely distributed throughout the body, and are involved in diverse physiological processes during both pre- and postnatal life. Like BMP-7, BMP-4 is involved in the development and maintenance of bone and cartilage. Reduced expression of BMP-4 is associated with a number of bone diseases, including the heritable disorder Fibrodysplasia Ossificans Progressiva.
BMP-5
TGF-β family members are key modulators of cell proliferation, differentiation, matrix synthesis, and apoptosis. As implied by their name, BMPs initiate, promote, and regulate the development, growth, and remodeling of bone and cartilage. In addition to this role, BMPs are also involved in prenatal development and postnatal growth, remodeling, and maintenance of a variety of other tissues and organs. BMP-5 is expressed in the nervous system, lungs and liver. It is a known regulator for dendritic growth in sympathetic neurons. BMP-5 is a 454 amino acid precursor protein that is cleaved to release the biologically active C-terminal mature protein.
BMP-6
TGF-β family members are key modulators of cell proliferation, differentiation, matrix synthesis, and apoptosis. As implied by their name, BMPs initiate, promote, and regulate the development, growth, and remodeling of bone and cartilage. In addition to this role, BMPs are also involved in prenatal development and postnatal growth, remodeling, and maintenance of a variety of other tissues and organs. Increasing evidence indicates that BMP-Smad signaling has a tumor suppressing activity, and that BMPs can inhibit tumor growth. BMP-6 is abnormally expressed in breast cancer cell lines, however, its function in promoting breast cancer development is unknown. The mature and functional form of BMP-6 is a homodimer of two identical 139 amino acid polypeptide chains linked by a single disulfide bond. Each monomer is expressed as the C-terminal part of a precursor polypeptide, which contains a 20 amino acid signal peptide and a 354 amino acid propeptide. This precursor undergoes intracellular dimerization, and upon secretion it is processed by a furin-type protease.
BMP-7
TGF-β family members are key modulators of cell proliferation, differentiation, matrix synthesis, and apoptosis. As implied by their name, BMPs initiate, promote, and regulate the development, growth, and remodeling of bone and cartilage. In addition to this role, BMPs are also involved in prenatal development and postnatal growth, remodeling, and maintenance of a variety of other tissues and organs. BMP-7, also known as osteogenic protein-1 or OP-1, is a potent bone inducing agent, which in the presence of an appropriate osteoconductive carrier (e.g. collagen sponge or synthetic hydroxyapatite) can be used in the treatment of bone defects. A bone-graft substitute, called OP-1TM implant, made of recombinant human BMP-7 associated with bovine bone-derived collagen, has recently been approved by the FDA as a device for treating critical-size bone fractures. The potential use of BMP-7 in dental reconstructive surgeries is currently under investigation.
Cardiotrophin-1 (CT-1)
CT-1 is a member of the IL-6 family of cytokines which also includes LIF, CNTF, OSM (Oncostatin M), IL-11, IL-6 and possibly NNT-1/BSF-3. CT-1 is a pleiotropic cytokine which is expressed in various tissues including the adult heart, skeletal muscle, ovary, colon, prostate and fetal lung and signals through the LIF receptor and the gp130 receptor subunit. CT-1 has the ability to induce cardiac myocyte hypertrophy, and enhances the survival of cardiomyocyte and different neuronal populations. Biologically active human CT-1 is synthesized as a 201 amino acid polypeptide lacking a hydrophobic N-terminal secretion signal sequence.
CDNF
CDNF is a secreted neurotrophic factor that is expressed in brain, neuronal and certain non-neuronal tissues. It has been shown to promote survival, growth and function of dopamine-specific neurons. CDNF and its structural homolog, MANF, each contain an N-terminal saposin-like lipid binding domain, and a carboxyl-terminal domain, which is not homologous to previously characterized protein structures. CDNF and MANF can prevent 6-OHDA-induced degeneration of dopaminergic neurons by triggering survival pathways in a rat experimental model of Parkinson's disease.
CNTF
CNTF is a potent neural factor that was originally characterized as a vital factor for the survival of chick ciliary neurons in vitro. CNTF is also important for the survival of other neural cell types, including primary sensory neurons, motor neurons, basal forebrain neurons and type 2 astrocytes. CNTF is highly conserved across species and exhibits cross-species bioactivity.
DKK-1
DKK-1 is a member of the DKK protein family which also includes DKK-2, DKK-3 and DKK-4. DKK-1 was originally identified as a Xenopus head-forming molecule that behaves as an antagonist for Wnt signaling. Subsequent studies have shown that DKK-1 and DKK-4 play an important regulatory role in the Wnt/β-catenin signaling pathway by forming inhibitory complexes with LDL receptor-related proteins 5 and 6 (LRP5 and LRP6), which are essential components of the Wnt/β-catenin signaling system. LRP5 and LRP6 are single-pass transmembrane proteins that appear to act as co-receptors for Wnt ligands involved in the Wnt/β-catenin signaling cascade. It has been suggested that by inhibiting Wnt/β-catenin signaling, which is essential for posterior patterning in vertebrates, DKK-1 permits anterior development. This notion is supported by the finding that mice deficient of DKK-1 expression lack head formation and die during embryogenesis.
DKK-2 / DKK-3
The dickkopf (DKK)-related protein family is comprised of four central members, DKK-1 - 4, along with the distantly-related DKK family member DKK-11 (Soggy), which is thought to be a descendent of an ancestral DKK-3 precursor due to its unique sequence homology to DKK-3 and no other DKK family member. DKK family members, with the exception of the divergent Soggy, share two conserved cysteine-rich domains and show very little sequence similarity outside of these domains. Playing an important regulatory role in vertebrate development through localized inhibition of Wnt-regulated processes, including anterior-posterior axial patterning, limb development, somitogenesis, and eye formation, DKKs have also been implicated post-developmentally in bone formation, bone disease, cancer, and neurodegenerative diseases. DKK proteins typically play an important regulatory role in the Wnt/β-catenin signaling pathway by forming inhibitory complexes with LDL receptor-related proteins 5 and 6 (LRP5 and LRP6), which are essential components of the Wnt/β-catenin signaling system. LRP5 and LRP6 are single-pass transmembrane proteins that appear to act as co-receptors for Wnt ligands involved in the Wnt/β-catenin signaling cascade. DKK-2 has been shown to both inhibit and enhance canonical Wnt signaling; enhancing Wnt signaling through direct high-affinity binding of DKK-2 to LRP6 during LRP6 overexpression, while inhibiting Wnt signaling and promoting LRP6 internalization through the formation of a ternary complex between DKK-2, LRP6, and Kremen-2. DKK-3 has been shown to potentiate, rather than inhibit, Wnt signaling through interactions with the high-affinity, transmembrane co-receptors Kremen-1 (Krm1) and Kremen-2 (Krm2).
sDLL-1
Human soluble DLL-1 comprises the extracellular signaling domain of DLL-1, a member of the Delta/Serrate/Lag-2 (DSL) family of single-pass type I trans-membrane proteins that serve as ligands for Notch receptors. It is expressed primarily in the heart, pancreas and epidermis. DLL-1 functions to specifically activate the Notch-1 and Notch-2 receptors. Proteolytic cleavage of DLL-1 produces a secreted extracellular domain, sDLL-1, that interacts with Notch receptors expressed on adjacent cells. Notch signaling plays an essential role in controlling cell fate decisions during prenatal development and postnatal stem cell renewal, and differentiation in many tissues. Human sDLL-1 blocks monocyte differentiation into macrophages, but permits differentiation into dendritic cells. In hematopoietic progenitor cells, hsDLL-1, suppresses differentiation into B-cells, while promoting differentiation into T-cells and NK cell precursors. In cell culture, human sDLL-1 has been shown to promote expansion of hematopoietic progenitor cells and suppress apoptosis by inhibiting differentiation. Overexpression of Notch receptors appears to inhibit differentiation in several mammalian cell lines, and increasing evidence suggests that Notch signaling is frequently downregulated in human malignancies. The human DLL-1 gene consists of a 528 amino acid extracellular domain with one DSL domain, eight EGF-like repeats, a 23 amino acid transmembrane domain, and a 155 amino acid cytoplasmic domain.
sDLL-4
Human sDLL-4 comprises the extracellular signaling domain of DLL, a member of a structurally-related family of single-pass type I trans-membrane proteins that serve as ligands for Notch receptors. DLL-4 functions to specifically activate the Notch-1 and Notch-4 receptors. The Notch signaling pathway regulates endothelial cell differentiation, proliferation and apoptosis, and is essential for the development, maintenance and remodeling of the vascular system. Targeted deletion of the DLL-4 gene in mice resulted in severe vascular defects and death before birth. Up-regulation of DLL-4 expression has been implicated in the vascular development of certain tumors. The human DLL-4 gene consists of a 503 amino acid extracellular domain with one DSL domain, eight EGF-like repeats, a 21 a.a. transmembrane domain, and a 135 a.a. cytoplasmic domain.
EGF
EGF is a potent growth factor that stimulates the proliferation of various epidermal and epithelial cells. Additionally, EGF has been shown to inhibit gastric secretion, and to be involved in wound healing. EGF signals through a receptor known as c-erbB, which is a class I tyrosine kinase receptor. This receptor also binds with TGF-α and VGF (vaccinia virus growth factor).
EGF Receptor (EGFR)
EGF Receptor (EGFR, ErbB1) is a transmembrane protein that exerts tyrosine kinase activity upon ligand-induced activation. EGFR can be activated by binding EGF, or at least six other structurally related protein ligands, including TGFα, HB-EGF, Betacellulin (BTC), Amphiregulin, Epiregulin, and Epigen. Upon activation, EGFR initiates a signaling cascade, which includes dimerization and internalization, tyrosine phosphorylation, DNA synthesis of target genes and, ultimately, cell proliferation. EGFR signaling plays a role in the growth and differentiation of normal cells, but elevated EGFR activity is correlated with the development and pathogenesis of certain cancers.
Eotaxin (CCL11)
Eotaxin is a CC chemokine that signals through the CCR3 receptor. It is produced by IFN-γ-stimulated endothelial cells and TNF-activated monocytes. Eotaxin selectively chemoattracts eosinophils and, along with Eotaxin-2 and Eotaxin-3, plays a key role in the regulation of eosinophil recruitment in the asthmatic lung and in allergic reactions.
EPO
Erythropoietin (EPO) is a glycoprotein hormone that is principally known for its role in erythropoiesis, where it is responsible for stimulating proliferation and differentiation of erythroid progenitor cells. The differentiation of CFU-E (Colony Forming Unit-Erythroid) cells into erythrocytes can only be accomplished in the presence of EPO. Physiological levels of EPO in adult mammals are maintained primarily by the kidneys, whereas levels in fetal or neonatal mammals are maintained by the liver. EPO also can exert various non-hematopoietic activities, including vascularization and proliferation of smooth muscle, neural protection during hypoxia, and stimulation of certain B cells.
FGF Family
Proteins of the FGF superfamily of growth factors manifest only a modest degree of primary sequence homology, yet share the ability to signal through one or more of four tyrosine kinase receptors called FGFR1 through FGFR4. The FGFs play a central role during prenatal development, postnatal growth and regeneration of a variety of tissues, by promoting cellular proliferation and differentiation. All members of the FGF superfamily bind, with varying degrees of affinity to heparin sulfate proteoglycans, which serve as extracellular storage sites and in some cases appear to be involved in the activation of the FGF receptors.
FGF-acidic (FGF-1)
FGF-acidic is one of 23 known members of the FGF family. Proteins of this family play a central role during prenatal development, postnatal growth and regeneration of a variety of tissues, by promoting cellular proliferation and differentiation. FGF-acidic is a non-glycosylated heparin binding growth factor that is expressed in the brain, kidney, retina, smooth muscle cells, bone matrix, osteoblasts, astrocytes and endothelial cells. FGF-acidic has the ability to signal through all the FGF receptors.
FGF-basic (FGF-2)
FGF-basic is one of 23 known members of the FGF family. Proteins of this family play a central role during prenatal development, postnatal growth and regeneration of a variety of tissues, by promoting cellular proliferation and differentiation. FGF-basic is a non-glycosylated, heparin-binding growth factor that is expressed in the brain, pituitary, kidney, retina, bone, testis, adrenal gland, liver, monocytes, epithelial cells and endothelial cells. FGF-basic signals through FGFR 1b, 1c, 2c, 3c and 4.
FGF-4
FGF-4 is a heparin-binding growth factor that is a member of the FGF family. Proteins of this family play a central role during prenatal development, postnatal growth and regeneration of a variety of tissues, by promoting cellular proliferation and differentiation. FGF-4 signals through the FGFR 1c, 2c, 3c, and 4.
FGF-5
FGF-5 is a secreted, heparin-binding growth factor that belongs to the FGF family. Proteins of this family play a central role during prenatal development, postnatal growth and regeneration of a variety of tissues, by promoting cellular proliferation and differentiation. FGF-5 binds to FGFR 1c and 2c, and plays a regulatory role in the hair growth cycle.
FGF-6
FGF-6 is a secreted, heparin-binding growth factor that is a member of the FGF family. Proteins of this family play a central role during prenatal development, postnatal growth and regeneration of a variety of tissues, by promoting cellular proliferation and differentiation. FGF-6 is expressed in leukemia cell lines with platelet megakaryocytic differentiation potential. It signals through FGFR 1c, 2c, and 4.
FGF-8 / FGF-8b
FGF-8 is a heparin-binding growth factor belonging to the FGF family. Proteins of this family play a central role during prenatal development, postnatal growth and regeneration of a variety of tissues, by promoting cellular proliferation and differentiation. There are 4 known alternate spliced forms of FGF8; FGF-8a, FGF-8b, FGF-8e and FGF-8f. The human and murine FGF-8a and b are identical, unlike human and mouse FGF-8e and f, which are 98% identical. FGF-8 targets mammary carcinoma cells and other cells expressing the FGF receptors.
FGF-9
FGF-9 is a heparin-binding growth factor that belongs to the FGF family. Proteins of this family play a central role during prenatal development, postnatal growth and regeneration of a variety of tissues, by promoting cellular proliferation and differentiation. FGF-9 targets glial cells, astrocytes cells and other cells that express the FGFR 1c, 2c, 3b, 3c, and 4.
FGF-10
FGF-10 is a heparin-binding growth factor that belongs to the FGF family. Proteins of this family play a central role during prenatal development, postnatal growth and regeneration of a variety of tissues, by promoting cellular proliferation and differentiation. FGF-10 is most related to KGF/FGF-7, and is expressed during the development and, preferentially, in adult lungs. It signals through the FGFR 2b.
FGF-16
FGF-16 is a heparin-binding growth factor that is a member of the FGF family. Proteins of this family play a central role during prenatal development, postnatal growth and regeneration of a variety of tissues, by promoting cellular proliferation and differentiation. FGF-16 signals through FGFR 2c and 3c. FGF-16 plays a role in the development of the central nervous system.
FGF-17
FGF-17 is a heparin-binding growth factor that is a member of the FGF family. Proteins of this family play a central role during prenatal development, postnatal growth and regeneration of a variety of tissues, by promoting cellular proliferation and differentiation. FGF-17 signals through the FGFR 1c, 2c, 3c, and 4. FGF-17 promotes the induction and patterning of brain development during embryogenesis.
FGF-18
FGF-18 is a heparin-binding growth factor that belongs to the FGF family. Proteins of this family play a central role during prenatal development, postnatal growth and regeneration of a variety of tissues, by promoting cellular proliferation and differentiation. FGF-18 is an essential regulator of long bone and calvarial development. FGF-18 signals through FGFR 1c, 2c, 3c, and 4.
FGF-19
The FGF family plays central roles during prenatal development and postnatal growth and regeneration of a variety of tissues, by promoting cellular proliferation and differentiation. FGF-19, a member of the FGF family, is a high-affinity heparin-dependent ligand for FGFR4. FGF-19 is expressed during brain development and embryogenesis.
FGF-20
FGF-20 is a secreted, heparin-binding growth factor that is a member of the FGF family. Proteins of this family play a central role during prenatal development, postnatal growth and regeneration of a variety of tissues, by promoting cellular proliferation and differentiation. FGF-20 signals through the FGFR 2c and 3c, and is expressed during limb and brain development.
FGF-21
FGF-21 is a secreted growth factor that is a member of the FGF family. Proteins of this family play a central role during prenatal development, postnatal growth and regeneration of a variety of tissues, by promoting cellular proliferation and differentiation. FGF-21, in the presence of β-Klotho as a protein cofactor, signals through the FGFR 1c and 4 receptors, and stimulates insulin-independent glucose uptake by adipocytes.
FGF-23
The FGF family plays a central role during prenatal development, postnatal growth and regeneration of a variety of tissues, by promoting cellular proliferation and differentiation. FGF-23, FGF-21 and FGF-19 constitute an atypical FGF subfamily whose ligands act as circulating hormones and require the participation of a Klotho protein as a co-receptor for their signaling. FGF-23 is a bone-derived hormone that acts in the kidney to regulate phosphate homeostasis and vitamin D metabolism. The signaling receptor for FGF-23, a Klotho-FGFR1 (IIIc) complex, is an essential regulator of the renal sodium phosphate co-transporter and key vitamin D-metabolizing enzymes CYP27B1 and CYP24A1.
FGFR1a / FGFR2a / FGFR3
The FGF family plays a central role during prenatal development and postnatal growth, and the regeneration of a variety of tissues, by promoting cellular proliferation and differentiation. The FGF ligands bind to a family of type I transmembrane tyrosine kinase receptors, which leads to dimerization and activation by sequential autophosphorylation of specific tyrosine residues. Four genes encoding structurally related FGF receptors (FGFR-1 to -4) are known. Alternative splicing of the mRNAs generates numerous forms of FGFR-1 to -3. Alternate forms of FGF receptors can exhibit different specificities with respect to ligand binding. For example, the form designated as FGFR1a (IIc) interacts predominantly with FGF-acidic (FGF1) and FGF-basic (FGF2). A frequent splicing event involving FGFR-1 and -2 results in receptors containing all three Ig domains, referred to as the alpha isoform, or only IgII and IgIII, referred to as the beta isoform. Only the alpha isoform has been identified for FGFR-3 and FGFR-4. Additional splicing events for FGFR-1 to -3, involving the C-terminal half of the IgIII domain encoded by two mutually exclusive alternative exons, generate FGF receptors with alternative IgIII domains (IIIb and IIIc).
Fractalkine (CX3CL1)
Fractalkine is a CX3CL chemokine that signals through the CX3CR1 receptor. Fractalkine has been shown to chemoattract monocytes, microglia cells and NK cells. Fractalkine is, at this time, the only CXC3C chemokine that contains three amino acid residues between the first and second cysteine residues of the chemokine domain. The Fractalkine gene encodes for a 397 amino acid precursor protein containing a 24 amino acid signal sequence, a chemokine domain, and a ìmucin-like stalkî sequence, which is followed by the transmembrane domain containing approximately 20 amino acids, and a C-terminal cytoplasmic domain. The extracellular chemokine domain contains 76 amino acid residues, including the four conserved cysteine residues found in other chemokines.
Furin
Proteases (also called Proteolytic Enzymes, Peptidases, or Proteinases) are enzymes that hydrolyze the amide bonds within proteins or peptides. Most proteases act in a specific manner, hydrolyzing bonds at, or adjacent to specific residues or a specific sequence of residues contained within the substrate protein or peptide. Proteases play an important role in most diseases and biological processes, including prenatal and postnatal development, reproduction, signal transduction, the immune response, various autoimmune and degenerative diseases, and cancer. They are also an important research tool, frequently used in the analysis and production of proteins. Furin is a calcium-dependent serine endoprotease that processes numerous proproteins of different secretory pathways into their mature forms by cleaving at the carboxyl side of the recognition sequence, R-Xaa-(K/R)-R, where Xaa can be any amino acid.
Galectin-1
Lectins, of either plant or animal origin, are carbohydrate-binding proteins that interact with glycoprotein and glycolipids on the surface of animal cells. The Galectins are lectins that recognize and interact with beta-galactoside moieties. Galectin-1 is an animal lectin that has been shown to interact with CD3, CD4, and CD45. It induces apoptosis of activated T-cells and T-leukemia cell lines, and inhibits the protein phosphatase activity of CD45.
Galectin-3
Lectins, of either plant or animal origin, are carbohydrate-binding proteins that interact with glycoproteins and glycolipids on the surface of animal cells. The Galectins are lectins that recognize and interact with β-galactoside moieties. Galectin-3 regulates a number of biological processes, including embryogenesis, inflammatory responses, cell progression and metastasis. Galectin-3 is normally expressed in epithelia of a variety of tissues, including colon and endometrium, and in various inflammatory cells, including macrophages. Galectin-3 can function intracellularly, controlling the cell cycle and preventing T-cell apoptosis, and also extracellularly, by activating various cells, including monocytes/macrophages, mast cells, neutrophils, and lymphocytes. Expression of Galectin-3 is affected by neoplastic transformation, being up-regulated in certain types of lymphomas, and in thyroid and hepatic carcinomas. Conversely, it is down-regulated in other cancers such as colon, breast, ovarian, and uterine.
GDF-2
GDF-2 belongs to the TGF-β cytokine family, whose members play an important role during prenatal development and postnatal growth, and the remodeling and maintenance of a variety of tissues and organs. GDF-2 is expressed mainly in non-parenchymal cells of the liver, but is also found in other various cells and tissues. GDF-2 can signal through the ALK1 receptor, and has been implicated in a number of physiologic events including the regulation of the hepatic reticuloendothelial system, glucose homeostasis, iron homeostasis, and the inhibition of angiogenesis.
GDF-7
GDF-7 belongs to the TGF-β superfamily of growth and differentiation factors. It is expressed selectively by roof plate cells that are located in the developing embryonic central nervous system, and has been shown to influence the neuronal identity of cells within the central nervous system. GDF-7 has also been implicated in the formation, maintenance, and repair of certain cartilage and ligament tissues.
GDNF
GDNF is a disulfide-linked, homodimeric neurotrophic factor structurally related to Artemin, Neurturin and Persephin. These proteins belong to the cysteine-knot superfamily of growth factors that assume stable dimeric protein structures. GDNF signals through a multicomponent receptor system, composed of a RET and one of the four GFRα (α1-α4) receptors. GDNF specifically promotes dopamine uptake and survival, and morphological differentiation of midbrain neurons. Using a Parkinson's disease mouse model, GDNF has been shown to improve conditions such as bradykinesia, rigidity, and postural instability. The functional human GDNF ligand is a disulfide-linked homodimer consisting of two 15 kDa polypeptide chains called monomers. Each monomer contains seven conserved cysteine residues, including Cys-101, which is used for inter-chain disulfide bridging, and others that are involved in the intramolecular ring formation known as the cysteine knot configuration.
GMF-β
GMF-β is a brain-specific protein that belongs to the actin-binding proteins ADF structural family. GMF-β appears to play a role in the differentiation, maintenance, and regeneration of the nervous system. It also supports the progression of certain auto-immune diseases, possibly through its ability to induce the production and secretion of various pro-inflammatory cytokines.
Gremlin-1
Gremlin-1 (isoform-1) belongs to a group of diffusible proteins that bind to ligands of the TGF-β family and regulate their activity by inhibiting their access to signaling receptors. The interplay between TGF-β ligands and their natural antagonists has major biological significance during development processes, in which cellular response can vary considerably depending upon the local concentration of the signaling molecule. Gremlin-1 is highly expressed in the small intestine, fetal brain, and colon; and is expressed at lower levels in the brain, prostate, pancreas, and in skeletal muscle. Gremlin-1 regulates multiple functions in early development by specifically binding to, and inhibiting the function of, BMP-2, -4, and -7. It also plays a role in carcinogenesis and kidney branching morphogenesis.
GRO/MGSA (CXCL1) / GROβ (CXCL2) / GROγ (CXCL3)
All three isoforms of GRO are CXC chemokines that can signal through the CXCR1 or CXCR2 receptors. The GRO proteins chemoattract and activate neutrophils and basophils. GRO/MGSA also stimulates mitogenesis in certain human melanoma cells.
HB-EGF
HB-EGF is an EGF-related growth factor that signals through the EGF receptor, and stimulates the proliferation of smooth muscle cells (SMC), fibroblasts, epithelial cells, and keratinocytes. HB-EGF is expressed in numerous cell types and tissues, including vascular endothelial cells, and vascular SMC, macrophages, skeletal muscle, keratinocytes, and certain tumor cells. The ability of HB-EGF to specifically bind heparin and heparin sulfate proteoglycans is distinct from other EGF-like molecules, and may be related to the enhanced mitogenic activity, relative to EGF, that HB-EGF exerts on smooth muscle cells. The human HB-EGF gene encodes a 208 amino acid transmembrane protein, which can be proteolytically cleaved to produce soluble HB-EGF.
Heregulinβ-1 / Neuregulin
Neuregulin/Heregulin is a family of structurally related polypeptide growth factors derived from alternatively spliced genes (NRG1, NRG2, NRG3 and NRG4). To date, there are over 14 soluble and transmembrane proteins derived from the NRG1 gene. Proteolytic processing of the extracellular domain of the transmembrane NRG1 isoforms releases soluble growth factors. HRG1-β1 contains an Ig domain and an EGF-like domain; the latter is necessary for direct binding to receptor tyrosine kinases erb3 and erb4. This binding induces erb3 and erb4 heterodimerization with erb2, stimulating intrinsic kinase activity that leads to tyrosine phosphorylation. Although HRG1-β1's biological effects are still unclear, it has been found to promote motility and invasiveness of breast cancer cells, which may also involve up-regulation of expression and function of the autocrine motility-promoting factor (AMF).
HGF
HGF is a potent, mesenchymally-derived mitogen for mature parenchymal hepatocytes, and acts as a growth factor for a broad spectrum of tissues and cell types. HGF signals through a transmembrane tyrosine kinase receptor known as MET. Activities of HGF include the induction of cell proliferation, motility, morphogenesis, inhibition of cell growth, and enhancement of neuron survival. HGF is a crucial mitogen for liver regeneration processes, especially after partial hepatectomy and other liver injuries. Human and murine HGF are cross-reactive. Human HGF is expressed as a linear, polypeptide-precursor glycoprotein containing 697 amino acid residues. Proteolytic processing of this precursor generates the biologically active heterodimeric form of HGF, which consists of two polypeptide chains (α-chain and β-chain) held together by a single disulfide bond resulting in formation of a biologically active heterodimer. The α-chain consists of 463 amino acid residues and four kringle domains. The β-chain consists of 234 amino acid residues.
I-309 (CCL1)
I-309 is a CC chemokine that signals through the CCR8 receptor. It is secreted by T lymphocytes, monocytes and mast cells. I-309 chemoattracts monocytes and Th2 differentiated T-cells, but not neutrophils. Human I-309 is active on murine cells. Unlike some CC chemokines, whose biologically active form in solution is a non-disulfide-linked dimer, the biologically active form of I-309 is monomeric.
IFN-α / IFN-β
Proteins of this family play an important role in inducing non-specific resistance against a broad range of viral infections. They also affect cell proliferation and modulate immune responses. Produced by peripheral blood leukocytes and lymphoblastoid cells, IFN-α is an acid-stable molecule that signals through IFN-α/βR, which is also used by IFN-β. Both IFNs have similar anti-viral activity and regulate expression of MHC class I antigens. IFN-α contains four highly conserved cysteine residues that form two disulfide bonds, one of which is necessary for biological activity.
IFN-γ
IFN-γ is an acid-labile interferon produced by CD4 and CD8 T lymphocytes as well as activated NK cells. IFN-γ receptors are present in most immune cells, which respond to IFN-γ signaling by increasing the surface expression of class I MHC proteins. This promotes the presentation of antigen to T-helper (CD4+) cells. IFN-γ signaling in antigen-presenting cells, and antigen-recognizing B and T lymphocytes, regulates the antigen-specific phases of the immune response. Additionally, IFN-γ stimulates a number of lymphoid cell functions, including the anti-microbial and anti-tumor responses of macrophages, NK cells, and neutrophils. Human IFN-γ is species-specific and is biologically active only in human and primate cells.
IGF-I / IGF-I LR3 / IGF-II
The IGFs are mitogenic, polypeptide growth factors that stimulate the proliferation and survival of various cell types, including muscle, bone, and cartilage tissue in vitro. IGFs are predominantly produced by the liver, although a variety of tissues produce the IGFs at distinctive times. The IGFs belong to the Insulin gene family, which also contains insulin and relaxin. The IGFs are similar to insulin by structure and function, but have a much higher growth-promoting activity than insulin. IGF-II expression is influenced by placenta lactogen, while IGF-I expression is regulated by growth hormone. Both IGF-I and IGF-II signal through the tyrosine kinase type I receptor (IGF-IR), but IGF-II can also signal through the IGF-II/Mannose-6-phosphate receptor. Mature IGFs are generated by proteolytic processing of inactive precursor proteins, which contain N-terminal and C-terminal propeptide regions. Recombinant human IGF-I and IGF-II are globular proteins containing 70 and 67 amino acids, respectively, and 3 intra-molecular disulfide bonds. IGF-I LR3 is a recombinant analog of human IGF-I comprised of the complete IGF-I sequence, with an Arginine substitution for the third position Glutamic acid, and a 13 amino acid length N terminus peptide extension. Specifically engineered for higher biological potency in vitro, IGF-I LR3 has an increased half-life and a binding aversion to native proteins within the body that make it ideal for both research and large-scale culturing.
IGF-BP4
IGF-BPs control the distribution, function and activity of IGFs in various cell tissues and body fluids. IGF-BP4 is the major IGF-BP produced by osteoblasts, and is found in the epidermis, ovarian follicles, and other tissues. IGF-BP4 inhibits the activity of IGF-I and IGF-II by binding in a manner that results in the formation of complexes with reduced ability to signal through cell surface IGF receptors. IGF-BP4 can inhibit the growth of chick pelvis cartilage and HT29 colon adenocarcinoma cells by blocking the mitogenic actions of IGFs, and has also been shown to reduce colony formation by colorectal cancer cells via an IGF-independent pathway. The biological effects of IGF-BP4 can be regulated by Pregnancy Associated Plasma Protein A (PAPP-A), which reduces IGF-BP4/IGF binding affinity by proteolytically cleaving IGF-BP4. The modulation of IGF-BP4 activity by PAPP-A is an important component in the regulation of ovarian folliculogenesis and in the growth inhibition of responding ovarian cancer cells.
IGF-BP6
IGF-BPs control the distribution, function and activity of IGFs in various cell tissues and body fluids. IGF-BP6, which specifically inhibits IGF-II actions, is produced by bone cells and is the major IGF-BP present in cerebrospinal fluid. IGF-BP6 has been shown to inhibit IGF-II-dependent cancers such as neuroblastoma, colon cancer and rhabdomyosarcoma.
IL-1α / IL-1β
IL-1α is a non-secreted, proinflammatory cytokine produced in a variety of cells, including monocytes, tissue macrophages, keratinocytes, and other epithelial cells. Both IL-1α and IL-1β bind to the same receptor and have similar, if not identical, biological properties. These cytokines have a broad range of activities including the stimulation of thymocyte proliferation by inducing IL-2 release, B-cell maturation and proliferation, mitogenic FGF-like activity and the release of prostaglandin and collagenase from synovial cells. However, whereas IL-1β is a secreted cytokine, IL-1α is predominantly a cell-associated cytokine.
IL-1 Receptor Antagonist (IL-1RA)
Interleukin-1 receptor antagonist (IL-1RA) is a naturally-occurring, inflammatory-inhibitor protein. It inhibits the activity of IL-1α and IL-1β by competitively blocking their binding to type I and type II receptors. IL-1RA is produced by corneal epithelial cells, monocytes, neutrophils, macrophages, and fibroblasts. Therapeutically, IL-1RA may help in the treatment of sepsis, cachexia, rheumatoid arthritis, chronic myelogenous leukemia, asthma, psoriasis, and inflammatory bowel disease.
IL-2
IL-2 is a powerful immunoregulatory lymphokine produced by T-cells in response to antigenic or mitogenic stimulation. IL-2/IL-2R signaling is required for T-cell proliferation and other fundamental functions that are essential for the immune response. IL-2 stimulates growth and differentiation of B-cells, NK cells, lymphokine-activated killer cells, monocytes, macrophages and oligodendrocytes.
IL-2 Receptor-α
The IL-2 receptor system consists of three non-covalently linked subunits termed IL-2Rα, IL-2Rβ, and IL-2Rγ. The IL-2Rα is a type I transmembrane protein consisting of a 219 amino acid extracellular domain, a 19 amino acid transmembrane domain and a 13 amino acid intracellular domain, which is not involved in the transduction of IL-2 signals. Proteolytic processing of IL-2Rα releases the entire extracellular domain of IL-2Rα, thereby generating a 219 amino acid soluble protein called soluble IL-2Rα (sIL-2Rα). The homodimeric form binds IL-2 (KD=10mM) and facilitates IL-2 signaling. The secreted sIL-2Rα is expressed on leukemia cells, lymphoma cells, and newly activated T and B cells, as well as on approximately 10% of NK cells.
IL-6
IL-6 is a pleiotropic cytokine that plays an important role in host defense by regulating immune and inflammatory responses. Produced by T cells, monocytes, fibroblasts, endothelial cells and keratinocytes, IL-6 has diverse biological functions. It stimulates B cell differentiation and antibody production, synergizes with IL-3 in megakaryocyte development and platelet production, induces expression of hepatic acute-phase proteins, and regulates bone metabolism. IL-6 signals through the IL-6 receptor system that consists of two chains, IL-6Rα and gp130. Murine IL-6 is inactive on human cells, while both human and murine are equally active on murine cells.
IL-6 Receptor-α
IL-6 mediates its biological effects through the type I IL-6 receptor system that consists of two chains, IL-6Rα and gp130. While the IL-6Rα chain is the binding component specific to IL-6, the gp130 chain only transmits signals of IL-6 when bound to IL-6Rα. The gp130 can also transmit signals from LIF, OSM, CNTF, IL-11 and CT-1 in conjunction with other receptor subunits. The low-affinity binding site for IL-6 is composed of IL-6Rα alone. IL-6Rα is expressed in a wide range of cells, including T cells, fibroblasts and macrophages. Soluble IL-6Rα, which consists of only the extracellular domain of the IL-6Rα chain, acts as an agonist of IL-6 activity at low concentrations.
IL-8 (CXCL8)
IL-8 is a proinflammatory CXC chemokine that can signal through the CXCR1 and CXCR2 receptors. It is secreted by monocytes and endothelial cells. IL-8 chemoattracts and activates neutrophils.
IL-10
IL-10 is an immunosuppressive cytokine produced by a variety of mammalian cell types including macrophages, monocytes, T cells, B cells and keratinocytes. IL-10 inhibits the expression of proinflammatory cytokines such as IL-1 and TNF-α. Like IL-4, IL-10 enhances humoral immune responses and attenuates cell-mediated immune reactions. Human IL-10 is active on murine cells, but murine IL-10 is inactive on human cells.
IL-11
IL-11 is a multifunctional cytokine produced by stromal cells, such as fibroblasts, epithelial cells and osteoclasts. It is expressed in a wide variety of tissues, including thymus, lung, bone, connective tissue and central nervous system. IL-11 plays an important regulatory role in hematopoiesis by stimulating growth of myeloid, erythroid and megakaryocyte progenitor cells. It also regulates bone metabolism, inhibits production of proinflammatory cytokines, and protects against gastromucosal injury.
IL-12
IL-12 is a potent regulator of cell-mediated immune responses and it induces IFN-γ production by NK and T cells. It is produced by activated monocytes/macrophage cells, B lymphocytes and connective tissue-type mast cells. Among its biological activities, IL-12 promotes the growth and activity of activated NK, CD4+ and CD8+ cells, and induces the development of IFN-γ-producing Th1 cells.
IL-15
IL-15 is an immunomodulating cytokine that stimulates the proliferation of T lymphocytes and shares many biological properties with IL-2. IL-15 exerts its biological activities primarily on T cells. It is also essential in the development, survival and activation of NK cells. Increased expression of IL-15 has been linked to rheumatoid arthritis, inflammatory bowel disease, and diseases affiliated with retroviruses HIV and HTLV-I. Human IL-15 is biologically active on mouse cells as measured by the dose-dependent stimulation of the proliferation of mouse CTLL cells.
IL-23
IL-23 is a proinflammatory, heterodimeric protein composed of two subunits: a unique p19 subunit, and a p40 subunit that is shared with IL-12. IL-23 is secreted by activated dendritic cells and macrophages, and signals though a receptor comprised of IL-23R complexed with IL-12Rβ2. IL-23 has been shown to enhance proliferation of memory T cells. It also stimulates the production of IFN-γ in NK cells, induces IL-17 production, and drives Th17-mediated responses.
IL-36 Receptor Antagonist (IL-36RA)
The IL-1 family is comprised of 11 structurally related ligands, including the recently re-named IL-36RA (IL-1F5), IL-36α (IL-1F6), IL-36β (IL-1F8), and IL-36γ (IL-1F9). The interaction of IL-36 ligands with the IL-1Rrp2 receptor (IL-1R6) can induce various activities, including dendritic cell maturation and activation. IL-36RA can antagonize the NF-κB signaling induced by either IL-36α, β or γ by binding to the IL-1Rrp2 receptor in a manner that prevents the initiation of functional signaling.
IL-36α (IL-1F6)
The IL-1 family is comprised of 11 structurally related ligands, including recently re-named IL-36α (IL-1F6), β (IL-1F8) and γ (IL-1F9). IL-36α is highly expressed in psoriatic plaques and in tissues containing epithelial cells. IL-36α signals through the IL-1Rrp2 (IL-1R6) receptor, which is primarily expressed on certain dendritic cells. The interaction of the IL-1Rrp2 receptor with IL-36 ligands induces dendritic cell maturation and activation. IL-36α also functions as an agonist of NF-κB, and can stimulate the inflammatory response in bronchial epithelial cells.
IL-36β (IL-1F8)
The IL-1 family is comprised of 11 structurally related ligands, including recently re-named IL-36α (IL-1F6), β (IL-1F8) and γ (IL-1F9). IL-36β is highly expressed in psoriatic plaques, and at lower levels in various other tissues. IL-36β signals through the IL-1Rrp2 (IL-1R6) receptor, which is primarily expressed on certain dendritic cells. The interaction of the IL-1Rrp2 receptor with IL-36 ligands induces dendritic cell maturation and activation. IL-36β also functions as an agonist of NF-κB, and stimulates the production of pro-inflammatory proteins, including IL-6, IL-8, BD-2, and BD-3.
IL-36γ(IL-1F9)
The IL-1 family is comprised of 11 structurally related ligands, including recently re-named IL-36α (IL-1F6), β (IL- 1F8) and γ (IL-1F9). IL-36γ is highly expressed in psoriatic plaques and in tissues containing epithelial cells. IL-36γ signals through the IL-1Rrp2 (IL-1R6) receptor, which is primarily expressed on certain dendritic cells. The interaction of the IL-1Rrp2 receptor with IL-36 ligands induces dendritic cell maturation and activation. IL-36γ also functions as an agonist of NF-κB, and can stimulate the inflammatory response in bronchial epithelial cells.
IL-37 (IL-1F7)
The IL-1 family is comprised of 11 structurally related ligands, including recently re-named IL-37 (IL-1F7), which acts as a modulator of the immune response. Reduction of IL-37 synthesis in PBMCs leads to increased production of pro-inflammatory cytokines including IL-1 alpha, IL-1 beta, IL-6 and TNF-alpha. The role of IL-37 as an inhibitor of the innate inflammatory response is also corroborated by the observation that it is highly expressed in synovial tissue from patients with rheumatoid arthritis. Full length IL-37 resides primarily in the cytoplasm, but after activation through cleavage by CASP1, it can translocate to the nucleus where it exerts its activity by direct interaction with SMAD3.
INSL5 / INSL7 Hybrid
The Insulin/Relaxin structural family of proteins consists of at least seven members, each containing an insulin-like heterodimeric structure of two polypeptide chains held together by two inter-chain disulfide bonds. The INSL5/INSL7 hybrid contains the β-chain of INSL7 (Relaxin-3), linked to the α-chain of INSL5. The INSL5/INSL7 hybrid protein can specifically activate the G protein-coupled receptors, GPCR135 and GPCR142, but unlike Relaxin-3, does not induce significant activation of the LGR7 receptor.
IP-10 (CXCL10)
IP-10 is a CXC chemokine that signals through the CXCR3 receptor. IP-10 selectively chemoattracts Th1 lymphocytes and monocytes, and inhibits cytokine-stimulated hematopoietic progenitor cell proliferation. Additionally, it is angiostatic and mitogenic for vascular smooth muscle cells.
Leptin
Encoded by the ob (obese) gene, Leptin is an adipose-derived cytokine that suppresses appetite and increases thermogenesis. Leptin exerts its anorectic effect via signaling through a hypothalamic receptor termed OB-R. Leptin has been shown to reduce body weight, food consumption, and plasma glucose levels in various in vivo models.
Leptin Receptor
Encoded for, along with leptin, by the obese (ob) gene, leptin receptor is a single-transmembrane-domain protein of the Type I, or Class I, Cytokine Receptor Family. The full length isoform, OB-Rb, is highly expressed in hypothalamic neurons, T-cells, and the vascular endometrium, and is thought to be the only isoform capable of transducing intracellular signals. Isoform OB-Ra, which is widely distributed at varying levels of expression, demonstrates weak signal activity and has been implicated in the active transport of leptin across the blood-brain barrier. Through ligand-binding with leptin receptor and the subsequent JAK2/STAT3 signaling cascade, the adipose-derived cytokine leptin functions to suppress appetite and increase thermogenesis. Leptin and leptin receptor have also, more recently, been implicated in the regulation or immune function, reproduction, glucose homeostasis, bone metabolism, wound healing, hematopoiesis, and angiogenesis. Mutations of the ob gene, which can result in leptin resistance and the down-regulation of ligand and/or receptor expression, have been connected to obesity and hypothalamic pituitary function in various in vivo models, including human, mouse, and rat.
LIF
LIF is a pleiotrophic factor produced by multiple cell types, including T cells, myelomonocytic lineages, fibroblasts, liver, heart and melanoma. LIF promotes long-term maintenance of embryonic stem cells by suppressing spontaneous differentiation. Other activities include the stimulation of acute phase protein synthesis by hepatocytes, stimulation of differentiation of cholinergic nerves, and suppression of adipogenesis by inhibiting the lipoprotein lipase in adipocytes. While human LIF is active on mouse cells and is widely used in the maintenance of murine ESC to prevent spontaneous differentiation, mouse LIF is not active on human cells due to its inability to bind to the human LIF receptor.
MANF
MANF is a secreted neurotrophic factor that is expressed in brain, neuronal and certain non-neuronal tissues. It has been shown to promote the survival, growth and function of dopamine-specific neurons. MANF and its structural homolog CDNF each contain an N-terminal, saposin-like, lipid-binding domain, and a carboxyl-terminal domain that is not homologous to previously characterized protein structures. MANF and CDNF can prevent 6-OHDA-induced degeneration of dopaminergic neurons by triggering survival pathways in a rat experimental model of Parkinson's disease.
MCP-1 (CCL2) / MCP-2 (CCL8) / MCP-3 (CCL7) / MCP-4 (CCL13) / MCP-5 (CCL12)
The MCP proteins are members of the CC chemokine family that signal through CCR2 and, with the exception of MCP-1, other CCR receptors. The MCP proteins chemoattract and activate monocytes, activated T cells, basophils, NK cells, and immature dendritic cells. The MCP family cross-reacts across species.
Midkine
Midkine (MK) and its functionally-related protein pleiotrophin are heparin-binding neurotrophic factors that signal through the same receptor, known as anaplastic lymphoma kinase (ALK). MK plays an important regulatory role in epithelial-mesenchymal interactions during fetal development and in postnatal lung development. MK chemoattracts embryonic neurons, neutrophils and macrophages, and exerts angiogenic, growth and survival activities during tumorgenesis.
MIP-1α (CCL3) / MIP-1β (CCL4)
Both MIP-1α and MIP-1β are structurally and functionally related CC chemokines. They participate in the host response to invading bacterial, viral, parasite and fungal pathogens by regulating the trafficking and activation state of selected subgroups of inflammatory cells, (e.g. macrophages, lymphocytes and NK cells). While both MIP-1α and MIP-1β exert similar effects on monocytes, their effect on lymphocytes differ; with MIP-1α selectively attracting CD8+ lymphocytes and MIP-1β selectively attracting CD4+ lymphocytes. Additionally, MIP-1α and MIP-1β have also been shown to be potent chemoattractants for B-cells, eosinophils and dendritic cells. Both human and murine MIP-1α and MIP-1β are active on human and murine hematopoietic cells.
Nesfatin-1
Nesfatin-1 is a metabolic polypeptide encoded in the N-terminal region of the precursor protein, Nucleobindin2 (NUCB2). Originally identified as a hypothalamic neuropeptide, nesfatin-1 is also expressed in other areas of the brain, in pancreatic islets β-cells, gastric endocrine cells, and adipocytes. Nesfatin-1 suppresses food intake and can regulate energy metabolism in a Leptin independent manner.
Neuritin
Neuritin is a neurotrophic factor that is expressed in response to the induction of neuronal activity by NGF, BDNF, NT3, and other neural stimulators. It is expressed primarily in postmitotic-differentiating neurons of the developing nervous system, and in neuronal structures related to synaptic plasticity in the adult nervous system. Neuritin acts as a molecular mediator of neurite outgrowth, neuronal survival, and synaptic maturation.
Neuropoietin
Neuropoietin is a newly identified member of the IL-6 cytokine family. Members of this family, including IL-6, IL-11, oncostatin M, leukemia inhibitory factor (LIF), cardiotrophin-1 (CT-1), cardiotrophin-like cytokine, and CNTF, display a four-helix bundle structure, and signal through gp130-containing receptor complexes. Neuropoietin, which is predominantly expressed in neuroepithelia during embryonic life, acts through a receptor complex formed of a CNTF receptor-α component, gp 130, and LIF receptor. Like CNTF, it promotes the survival of embryonic motor neurons, and could increase the proliferation of neural precursor cells in the presence of EGF and FGF-2. Interestingly, the human neuropoietin gene has evolved toward a pseudogene, suggesting that alternative signaling via CNTF is an effective compensatory pathway.
Neuroserpin
Neuroserpin is an inhibitory serpin that is expressed predominantly in the central nervous system. Although the physiological target of neuroserpin is still unclear, cumulative evidence suggests that it plays an important role in controlling proteolytic degradation of extracellular matrix (ECM) during synaptogenesis, and the subsequent development of neuronal plasticity. In the adult brain, neuroserpin is secreted from the growth cones of neurons in areas where synaptic changes are associated with learning and memory (i.e. cerebral cortex, hippocampus, and amygdala.) The neuroprotective role of neuroserpin has been demonstrated in transgenic mice lacking neuroserpin expression. The deficiency of neuroserpin in these mice was associated with motor neuron disease characterized by axonal degradation. In humans, defects in neuroserpin, caused by point mutations in the neuroserpin gene, underlie a hereditary disorder called the familial encephalopathy with neuroserpin inclusion bodies (FENIB).
Neurturin
Neurturin is a disulfide-linked homodimer neurotrophic factor structurally related to GDNF, artemin, and persephin. These proteins belong to the cysteine-knot family of growth factors that assume stable dimeric structures. Neurturin signals through a multicomponent receptor system, composed of RET and one of four GFRα (α1-α4) receptors. Neurturin promotes the development and survival of sympathetic and sensory neurons by signaling through a receptor system composed of RET and GFRα2.
β-NGF
β-NGF is a neurotrophic factor structurally related to BDNF, NT-3 and NT-4. These proteins belong to the cysteine-knot family of growth factors that assume stable dimeric structures. β-NGF is a potent neurotrophic factor that signals through its receptor β-NGFR, and plays a crucial role in the development and preservation of the sensory and sympathetic nervous systems. β-NGF also acts as a growth and differentiation factor for B lymphocytes, and enhances B-cell survival.
NNT-1 / BCSF-3
NNT-1/BCSF-3 is a neurotrophic factor with B-cell-stimulating capabilities. Expressed in the lymph nodes and spleen, NNT-1/BCSF-3 activates glycoprotein 130 (gp130) and leukemia inhibitory factor receptor member β (LIFR-β) by binding and inducing the tyrosine phosphorylation of these receptors. In vitro, it supports the survival of chick embryo motor and sympathetic neurons. In mice, NNT-1/BCSF-3 induces serum amyloid A, causes body weight loss, and B-cell hyperplasia associated with increases in serum IgG and IgM.
Noggin
Noggin belongs to a group of diffusible proteins that bind to ligands of the TGF-β family, and regulate their activity by inhibiting their access to signaling receptors. The interplay between TGF-β ligands and their natural antagonists has major biological significance during development processes, in which cellular response can vary considerably depending upon the local concentration of the signaling molecule. Noggin was originally identified as a BMP-4 antagonist whose action was critical for proper formation of the head and other dorsal structures. Consequently, noggin has been shown to modulate the activities of other BMPs including BMP-2,-7,-13, and -14. Targeted deletion of Noggin in mice results in prenatal death, and a recessive phenotype displaying a severely malformed skeletal system. Conversely, transgenic mice over-expressing noggin in mature osteoblasts display impaired osteoblastic differentiation, reduced bone formation, and severe osteoporosis.
NT-3
NT-3 is a neurotrophic factor structurally related to β-NGF, BDNF, and NT-4. These proteins belong to the cysteine-knot family of growth factors that assume stable dimeric structures. NT-3 is expressed by neurons of the central nervous systems, and can signal through the trk receptors. NT-3 promotes the growth and survival of nerve and glial cells. The amino acid sequences of human, murine and rat NT-3 are identical.
NT-4
NT-4 is a neurotrophic factor structurally related to β-NGF, BDNF, and NT-3. These proteins belong to the cysteine-knot family of growth factors that assume stable dimeric structures. NT-4 is expressed in the prostate, thymus, placenta, and skeletal muscle. NT-4 can signal through the LNGFR and trkB receptors, and promotes the survival of peripheral sensory sympathetic neurons.
Oncostatin M
Oncostatin M (OSM) is a growth and differentiation factor that participates in the regulation of neurogenesis, osteogenesis and hematopoiesis. Produced by activated T cells, monocytes and Kaposi's sarcoma cells, OSM can exert both stimulatory and inhibitory effects on cell proliferation. It stimulates the proliferation of fibroblasts, smooth muscle cells and Kaposi's sarcoma cells, but inhibits the growth of some normal and tumor cell lines. It also promotes cytokine release (e.g. IL-6, GM-CSF and G-CSF) from endothelial cells, and enhances the expression of low-density lipoprotein receptors in hepatoma cells. OSM shares several structural and functional characteristics with LIF, IL-6, and CNTF.
PAI-1
Plasminogen Activator Inhibitor-1 (PAI-1, Serpin E1) is a member of the serpin family of serine protease inhibitors, and is the primary inhibitor of urokinase and tissue plasminogen activator (tPA). PAI-1 is expressed predominantly in adipose, liver and vascular tissues, but is also produced by certain tumor cells. Elevated levels of PAI-1 are associated with obesity, diabetes and cardiovascular disease, and increased production of PAI-1 is induced by various obesity-related factors, such as TNFα, glucose, insulin, and very-low-density lipoprotein. The obesity-related elevation of PAI-1 levels, along with the consequential deficiency in plasminogen activators, can lead directly to increased risk of thrombosis and other coronary diseases. Accordingly, PAI-1 has been implicated as an important molecular link between obesity and coronary disease. PAI-1 can also specifically bind vitronectin (VTN) to form a stable active complex with an increased circulatory half-life relative to free PAI-1.
PDGF-AA / PDGF-AB / PDGF-BB
PDGFs are disulfide-linked dimers consisting of two 12.0-13.5 kDa polypeptide chains, designated PDGF-A and PDGF-B chains. The three naturally occurring PDGFs, PDGF-AA, PDGF-BB and PDGF-AB, are potent mitogens for a variety of cell types, including smooth muscle cells, connective tissue cells, bone and cartilage cells, and some blood cells. The PDGFs are stored in platelet α-granules, and are released upon platelet activation. The PDGFs are involved in a number of biological processes, including hyperplasia, chemotaxis, embryonic neuron development, and respiratory tubule epithelial cell development. Two distinct signaling receptors used by PDGFs have been identified and named PDGFR-α and PDGFR-β. PDGFR-α is high-affinity receptor for each of the three PDGF forms. On the other hand, PDGFR-β interacts with only PDGF-BB and PDGF-AB.
PDGF-CC
The platelet-derived growth factor (PDGF) family of heparin-binding growth factors consists of five known members, denoted PDGF-AA, PDGF-BB, PDGF-AB, PDGF-CC and PDGF-DD. The mature and active form of these proteins, an anti-parallel disulfide-linked dimer of two 12-14 kDa polypeptide chains, is obtained through proteolytic processing of biologically inactive precursor proteins, which contain an N-terminal CUB domain and a PDGF/VEGF homologous domain. The PDGFs interact with two related protein tyrosine kinase receptors, PDGFR-α and PDGFR-β, and are potent mitogens for a variety of cell types, including smooth muscle cells, connective tissue cells, bone and cartilage cells, and certain tumor cells. They play an important role in a number of biological processes, including hyperplasia, chemotaxis, embryonic neuron development, and respiratory tubules epithelial cell development. Mature PDGFs are stored in platelet α-granules and are released upon platelet activation. PDGF-AA, -AB, -BB and -CC signal primarily through the PDGF-Rα receptor, whereas PDGF-DD interacts almost exclusively with the PDGF-Rβ receptor.
PEDF
PEDF is a noninhibitory serpin with neurotrophic, anti-angiogenic, and anti-tumorigenic properties. It is a 50 kDa glycoprotein produced and secreted in many tissues throughout the body. A major component of the anti-angiogenic action of PEDF is the induction of apoptosis in proliferating endothelial cells. In addition, PEDF is able to inhibit the activity of angiogenic factors, such as VEGF and FGF-2. The neuroprotective effects of PEDF are achieved through suppression of neuronal apoptosis induced by peroxide, glutamate, or other neurotoxins. The recent identification of a lipase-linked cell membrane receptor for PEDF (PEDF-R) that binds to PEDF with high affinity (Notari, I. et al. J Biol Chem., Vol. 281, 38022-38037) should facilitate further elucidation of the underlying mechanisms of this pluripotent serpin. To date, PEDF-R is the only signaling receptor known to be used by a serpin family member. The unique range of PEDF activities implicate it as a potential therapeutic agent for the treatment of vasculature-related neurodegenerative diseases, such as age-related macular degeneration (AMD) and proliferative diabetic retinopathy (PDR). PEDF also has the potential to be useful in the treatment of various angiogenesis-related diseases including a number of cancers.
Persephin
Persephin is a disulfide-linked, homodimeric, neurotrophic factor structurally related to GDNF, artemin, and neurturin. These proteins belong to the cysteine knot family of growth factors that assume stable dimeric structures. Persephin signals through a multicomponent receptor system, composed of RET and one of four GFR α (α1-α4) receptors. The GFRα4 was first identified in chicken, and was later shown to be the preferential binding subunit for persephin. Persephin promotes the survival of ventral midbrain dopaminergic neurons and motor neurons after sciatic nerve oxotomy, and, like GDNF, promotes ureteric bud branching. However, in contrast to GDNF and neurturin, persephin does not support the survival of peripheral neurons.
Pleiotrophin
Pleiotrophin and Midkine are structurally related heparin-binding neurotrophic factors, whose expression is developmentally regulated. The expression pattern of these neurotrophic factors suggests function in neurogenesis, cell migration, secondary organogenetic induction, and mesoderm epithelial interaction. The expression of PTN increases during the process of brain embryogenesis, and reaches maximum levels at time of birth. The physiological roles of PTN and Midkine are largely unknown, but these neurotrophins have been implicated in the pathogenesis of neuroblastomas.
PlGF-1
PlGF-1 is an angiogenic factor that belongs to the cysteine-knot superfamily of growth factors. PlGF-1 is expressed in placental tissues, the colon, and mammary carcinomas. It signals through the VEGFR-1/FLT1 receptor, and stimulates endothelial cell proliferation and migration.
Prolactin
Prolactin is a neuroendocrine hormone secreted by the pituitary gland. Its primary function is to promote and maintain lactation during pregnancy and suckling. In addition, prolactin plays an immunoegulatory role by stimulating the activities of ornithine decarboxylase and protein kinase C, which are important for the proliferation, differentiation, and function of lymphocytes.
RANTES (CCL5)
RANTES is a CC chemokine that can signal through the CCR1, CCR3, CCR5 and US28 (cytomegalovirus receptor) receptors. It is a chemoattractant towards monocytes, memory T cells (CD4+/CD45RO), basophils, and eosinophils. RANTES also has the capability to inhibit certain strains of HIV-1, HIV-2 and simian immunodeficiency virus (SIV).
Relaxin-3
Relaxin-3 is a secreted protein structurally related to insulin that is expressed primarily in the brain and central nervous system. Relaxin-3 has been identified as the ligand for the GPCR135 receptor, previously known as "somatostatin-like" or "angiotensin-like" peptide receptor, and has also been identified for binding specifically to the LGR7 receptor, previously identified as an "orphan" G protein-coupled receptor. Signaling by relaxin-3 through its target receptors is, most likely, part of a CNS processing system, activated in response to signaling by neuropeptides and other factors. Intracerebroventricular injections of relaxin-3 have been shown to cause a significant increase of food intake and body weight in Wistar rats.
ROR1
Receptor-tyrosine-kinase-like orphan receptor 1 (ROR1) is a tumor-associated, surface protein predominantly expressed during embryogenesis, where it is involved in organ morphogenesis, nervous system development, and neural progenitor cell maintenance and survival. Virtually absent from normal pediatric and adult tissues, with the exception of low-level expression in a subset of immature B-cell precursors known as hematogones and adipocytes, ROR1 is notably overexpressed, and considered a survival factor, in a number of B lymphoid and epithelial malignancies: most notably, but not exclusively, including chronic lymphocytic leukemia (CLL), mantle cell lymphoma (MCL), acute lymphoblastic leukemia (ALL), marginal zone lymphoma, lung adenocarcinoma. First identified during PCR-based cloning of a human neuroblastoma cell line in search of tyrosine kinases similar to tropomyosin-receptor-kinase (Trk) neurotropic receptors, ROR1, along with the related receptor tyrosine kinase (RTK) ROR2, was catalogued as an ìorphanî receptor due to the fact its related ligand remained elusive. Wnt-5a has since been suggested as a candidate ligand for ROR1, and ROR1 has been implicated to function as a pseudokinase, promoting proliferation and resistance to apoptosis in cancer cells through interaction with Wnt-5a, and TCL1-co-activation of AKT. ROR1 is expressed as a glycoprotein containing extracellular immunoglobulin (Ig)-like, Frizzled, and Kringle domains, as well as an intracellular region containing a tyrosine kinase domain.
R-Spondin-1
R-Spondin-1 (Rspo-1) belongs to the (Rspo) family of Wnt modulators. Currently, the family consists of four structurally related secreted ligands (Rspo 1-4), all containing the furin-like and thrombospondin structural domains. Rspo-1 is expressed in certain areas of the developing central nervous system, as well as in the adrenal glands, ovary, testis, thyroid, and trachea. Rspo can interact with the Frizzled/LRP6 receptor complex in a manner that stimulates the Wnt/β-catenin signaling pathway.
R-Spondin-2 / R-Spondin-3
The R-Spondin (Rspo) proteins belong to the Rspo family of Wnt modulators. Currently, the family consists of four structurally-related, secreted ligands (Rspo 1-4), all containing furin-like and thrombospondin structural domains. The Rspo proteins can interact with the Frizzled/LRP6 receptor complex in a manner that causes the stabilization, and resulting accumulation, of the intracellular signaling protein, β-catenin. This activity effectively activates and increases the subsequent nuclear signaling of β-catenin. R-Spondin can also bind to the previously discovered G-protein coupled receptors, LGR-4 and LGR-5. Rspo/β-catenin signaling can act as an inducer of the transformed phenotype, and can also regulate the proliferation and differentiation of certain stem cell populations.
SDF-1α (CXCL12) / SDF-1β (CXCL12)
SDF-1α and β are stromal-derived, CXC chemokines that signal through the CXCR4 receptor. SDF-1α and β chemoattract B and T cells, and have been shown to induce migration of CD34+ stem cells. Additionally, the SDF-1 proteins exert HIV-suppressive activity in cells expressing the CXCR4 receptor. Human and murine SDF-1 proteins act across species. SDF-1α and β contain the four highly conserved cysteine residues present in CXC chemokines. The mature SDF-1α protein is the result of alternative splicing of the SDF-1 gene and contains 68 amino acid residues. The mature SDF-1β protein, produced by an N-terminal truncation of two additional amino acids, after removal of the signal sequence, contains 72 amino acid residues.
Semaphorin 3A
Semaphorins are a large group of structurally-related, secreted, GPI-anchored, transmembrane, cell-signaling molecules. There are 8 major classifications of Semaphorins (the first seven ordered by number, 1-7, and the eighth designated V for virus), which are characterized by the existence of a conserved 500 amino acid SEMA domain at the amino terminus. Classes 3, 4, 6, and 7 are found in vertebrates only, whilst class 5 is found in both vertebrates and invertebrates. Each class is then divided into additional subgroups based on shared structural characteristics. Semaphorins primarily function as axon growth cone guidance factors during neuronal development. Semaphorin 3A acts as a chemo-repellent to axons, and an inhibitor of the growth of axons by signaling through receptors, Neuropilin-1 and Plexin-A.
Slit2-N
Slit2 is a member of the Slit family that signals through the Roundabout (Robo) receptor as a repellent for axon guidance and neuronal migration, and also acts as a chemoattractant to vascular endothelial cells and a chemotaxis inhibitor for leukocytes. Slit2 is expressed primarily in the fetal lung, kidney, and adult spinal cord, and to a lesser extent in the adult adrenal gland, thyroid and trachea. Slit2 is initially synthesized as a 1499 amino acid precursor, which is subsequently cleaved into N-terminal and C-terminal fragments, designated as Slit2-N and Slit2-C respectively. The neurodevelopment-related activities, as measured by the ability to repel olfactory bulb axons and to induce branching in dorsal root ganglia axons, are contained only in the N-terminal fragment.
Sonic Hedgehog (Shh)
Members of the Hedgehog (Hh) family are highly conserved proteins that are widely represented throughout the animal kingdom. The three known mammalian Hh proteins, Sonic (Shh), Desert (Dhh) and Indian (Ihh), are structurally related, and share a high degree of amino acid sequence identity (e.g. Shh and Ihh are 93% identical). The biologically active form of each Hh molecule is obtained by autocatalytic cleavage of their precursor proteins, and each corresponds to approximately one half of the N-terminal portion of the precursor molecule. Although Hh proteins have unique expression patterns and distinct biological roles within their respective regions of secretion, they use the same signaling pathway and can be substituted for one another in experimental systems.
TAFA-2
TAFA proteins are a newly discovered family of proteins, which are distantly related to MIP-1α, a member of the CC-chemokine family. TAFA mRNAs are highly expressed in specific brain regions. The biological function of TAFA-2 is still unknown.
TGF-α
TGF-α is an EGF-related polypeptide growth factor that signals through the EGF receptor, and stimulates the proliferation of a wide range of epidermal and epithelial cells. It is produced by monocytes, keratinocytes, and various tumor cells. TGF-α induces anchorage-independent transformation in cultured cells. Human, murine and rat TGF-α are cross-species reactive.
TGF-β1 / TGF-β2 / TGF-β3
The three mammalian isoforms of TGF-β, TGF-β1, β2, β3, signal through the same receptor and elicit similar biological responses. They are multifunctional cytokines that regulate cell proliferation, growth, differentiation and motility, as well as synthesis and deposition of the extracellular matrix. They are involved in various physiological processes including embryogenesis, tissue remodeling and wound healing. They are secreted predominantly as latent complexes, which are stored at the cell surface and in the extracellular matrix. The release of biologically active TGF-β isoform from a latent complex involves proteolytic processing of the complex and /or induction of conformational changes by proteins such as Thrombospondin-1.
TGF-β1: TGF-β1 is the most abundant isoform secreted by almost every cell type. It was originally identified for its ability to induce phenotypic transformation of fibroblasts, and recently it has been implicated in the formation of skin tumors.
TGF-β2: TGF-β2 has been shown to exert suppressive effects on IL-2-dependent T-cell growth, and may also have an autocrine function in enhancing tumor growth by suppressing immuno-surveillance of tumor development.
TGF-β3: The physiological role of TGF-β3 is still unknown, but its expression pattern suggests a role in the regulation of certain development processes.
TIMP-1
TIMP-1 is an extracellular inhibitor of MMPs, including MMP-1, -2, -3, -7, -8, -9, -10, -11, -12, -13, and -16. It belongs to the I35 (TIMP) family of irreversible protease inhibitors that function as key modulators of extracellular matrix degradation during tissue development and remodeling. TIMP-1 can also act through an MMP-independent mechanism to promote erythropoiesis by stimulating proliferation and differentiation of erythroid progenitors.
TNF-α (TNFSF1A)
TNF-α is a pleiotropic pro-inflammatory cytokine secreted by various cells, including adipocytes, activated monocytes, macrophages, B cells, T cells and fibroblasts. It belongs to the TNF family of ligands, and signals through two receptors, TNFR1 and TNFR2. TNF-α is cytotoxic to a wide variety of tumor cells, and is an essential factor in mediating the immune response against bacterial infections. TNF-α also plays a role in the induction of septic shock, autoimmune diseases, rheumatoid arthritis, inflammation, and diabetes. Human and murine TNF-α demonstrate significant cross-species reactivity. TNF-α exists in two forms; a type II transmembrane protein, and a mature soluble protein. The TNF-α transmembrane protein is proteolytically cleaved to yield a soluble, biologically active, 17 kDa TNF-α, which forms a non-covalently linked homotrimer in solution.
TNF-β (TNFSF1B)
TNF-β is a potent mediator of inflammatory and immune responses. It belongs to the TNF family of ligands, and signals through TNFR1 and TNFR2. TNF-β is produced by activated T and B lymphocytes, and has similar activities to TNF-α. Like TNF-α, TNF-β is involved in the regulation of various biological processes, including cell proliferation, differentiation, apoptosis, lipid metabolism, coagulation, and neurotransmission. TNF-β is secreted as a soluble polypeptide, but can form heterotrimers with lymphotoxin-β, which effectively anchors the TNF-β to the cell surface. TNF-β is cytotoxic to a wide range of tumor cells.
sTNF-Receptor Type I (TNFR1)
TNFRI belongs to the TNFR superfamily of transmembrane proteins, and is expressed in most cell types. Binding of either TNF-α or TNF-β to TNFRI initiates a signal transduction pathway that results in the activation of the transduction factor NF-κB, whose target genes are involved in the regulation of inflammatory responses, and, in certain cells induce apoptosis. Soluble TNF Receptor I (sTNFRI) is capable of inhibiting TNF-α and TNF-β activities by acting as a decoy receptor that serves as a sink for the TNF ligands. The human TNFRI gene encodes for a 455 amino acid type I transmembrane protein, which contains a 21 amino acid signal sequence, a 190 amino acid extracellular domain, a 23 amino acid transmembrane domain, and a 221 amino acid cytoplasmic domain.
sTNF-Receptor Type II (TNFR2)
TNFRII is a member of the TNFR family of transmembrane proteins, and is expressed in immune cells and certain endothelial cells. It is a high affinity receptor for TNF-α, but manifests a lower affinity to TNF-β. Signaling through this receptor regulates various biological processes, including cell proliferation, differentiation, apoptosis, lipid metabolism, coagulation, and neurotransmission. Soluble TNFRII is capable of inhibiting TNF-α-induced activities by acting as a decoy receptor. The human TNFRII gene encodes for a 461 amino acid type I transmembrane protein, which contains a 22 amino acid signal sequence, a 235 amino acid extracellular domain, a 30 amino acid transmembrane domain, and a 174 amino acid cytoplasmic domain.
TWEAK (TNFSF12)
TWEAK belongs to the TNF family of ligands, and signals through TWEAKR, also known as TNFRSF12A. TWEAK is expressed in a variety of tissues, including the adult heart, pancreas, skeletal muscle, small intestine, spleen and peripheral blood lymphocytes. TWEAK has the ability to induce NF-κB activation and chemokine secretion, and to exert an apoptotic activity in certain cells, such as HT-29 human adenocarcinoma cells when cultured in the presence of IFN-γ. TWEAK also promotes proliferation and migration of endothelial cells. The human TWEAK gene encodes for a 249 amino acid type II transmembrane protein, which contains a 21 amino acid cytoplasmic domain, a 21 amino acid transmembrane domain, and a 207 amino acid extracellular domain.
VEGF-A
VEGF is a potent growth and angiogenic cytokine. It stimulates proliferation and survival of endothelial cells, and promotes angiogenesis and vascular permeability. Expressed in vascularized tissues, VEGF plays a prominent role in normal and pathological angiogenesis. Substantial evidence implicates VEGF in the induction of tumor metastasis and intra-ocular neovascular syndromes. VEGF signals through three receptors; fms-like tyrosine kinase (flt-1), KDR gene product (the murine homolog of KDR is the flk-1 gene product) and the flt4 gene product.
VEGF-B
VEGF-B, a member of the VEGF family, is a potent growth and angiogenic cytokine. It promotes DNA synthesis in endothelial cells, helps regulate angiogenesis and vascular permeability, and inhibits apoptosis in certain smooth muscle cells and neurons. VEGF-B is expressed in all tissues except the liver. It forms cell surface-associated, disulfide-linked homodimers, and can form heterodimers with VEGF-A. There are two known isoforms, formed by alternative splicing, which have been designated VEGF-B167 and VEGF-B186. Both forms have identical amino-terminal sequences encoding a cysteine knot-like structural motif, but differ in their carboxyl-terminal domains. Both VEGF-B isoforms signal only through the VEGFR1 receptor.
VEGF-C
VEGF-C, a member of the VEGF/PDGF family of structurally related proteins, is a potent angiogenic cytokine. It promotes endothelial cell growth, promotes lymphangiogesis, and can also affect vascular permeability. VEGF-C is expressed in various tissues, but is not produced in peripheral blood lymphocytes. It forms cell surfaced-associated, non-covalent, disulfide-linked homodimers, and can bind and activate both VEGFR-2 (flk1) and VEGFR-3 (flt4) receptors. During embryogenesis, VEGF-C may play a role in the formation of the venous and lymphatic vascular systems. Both VEGF-C and VEGF-D are over-expressed in certain cancers, and the resulting elevated levels of VEGF-C or VEGF-D tend to correlate with increased lymphatic metastasis.
Visfatin
Visfatin is a 55 kDa protein produced and secreted primarily by white adipose tissue. Recently, visfatin was isolated from visceral fat deposits and shown to possess insulin-mimetic activity. Like insulin, visfatin exerts hypoglycemic effects by interacting with the insulin receptor. The binding affinity of visfatin for the insulin receptor is similar to that of insulin, but it does not compete with insulin, suggesting that the two proteins interact with different receptor sites. The circulating levels of visfatin are much lower than those of insulin and are not affected by feeding, implying that the hypoglycemic effect of visfatin may not be of physiological importance. The plasma visfatin levels, like those of leptin, correlate positively with the percent of body fat, and increase during the development of obesity. Another similarity between visfatin and leptin is that their amino acid sequence is highly conserved across different mammalian species and shows no homology to any known protein. Receptors for both leptin (Ob-R) and visfatin (i.e. the insulin receptor) are expressed by neurons within the arcuate nucleus of the hypothalamus, a brain area that plays a pivotal role in the regulation of energy metabolism. Although the metabolic function of visfatin is still unknown, it appears that this newly identified adipocytokine might play an important role, similar to that of leptin, in the regulation of body weight, i.e. as an afferent signal reflecting excess body fat. The PBEF gene encodes a polypeptide of 491 amino acid residues. The secreted form of this polypeptide, i.e. visfatin, contains 465 residues and lacks the first 26 N-terminal residues of the PBEF gene product. The 491-residue form has been shown to be a nicotinamide phosphoribosyltransferase, a cytosolic enzyme involved in NAD biosynthesis. The amino acid sequence of visfatin is highly conserved across different species and shows no homology to any known protein. It contains 5 cysteine residues, of which only two of them appear to be involved in disulfide bridge formation.
Vitronectin
Vitronectin is a secreted glycoprotein that is synthesized in the liver. It circulates primarily in monomeric form, but can undergo conformational change to a structure that forms disulfide-linked multimers. The multimeric vitronectin can efficiently bind to, and incorporate into, the extracellular matrix. Within the matrix, vitronectin can support cell adhesion through binding to various integrins and other proteoglycans. Additionally, recombinant vitronectin can function as a chemically-defined matrix component in human embryonic stem cell renewal media.
WISP-1
WISP-1 is a member of the CCN family of secreted, cysteine-rich regulatory proteins. It is expressed in the heart, kidney, lung, pancreas, placenta, ovary, small intestine and spleen. WISP-1 is a β-catenin-regulated protein that can contribute to tumorigenesis, and has also been shown to play a role in bone development and fracture repair.
Wnt-1
Wnt-1 is a secreted protein that signals through the Frizzled family of cell surface receptors, and is required for normal embryonic development. Wnt-1 activation induces a complex signaling cascade that ultimately leads to the increased expression of over fifty genes. An important component of Wnt-1 signaling is the stabilization, and resulting accumulation, of the intracellular signaling protein, β-catenin. Wnt signaling induces and maintains the transformed phenotype, and, in certain embryonic cell lines, supports self-renewal in the absence of significant differentiation. Elevated levels of Wnt proteins are associated with tumorigenesis, and are present in numerous human breast cancers. Mature human Wnt-1 is a glycosylated protein containing 343 amino acid residues.
Wnt-3a
Wnt-3a belongs to the Wnt family of signaling proteins that play a key role in maintaining the integrity of embryonic and adult tissues. Expression of Wnt-3a occurs primarily along the dorsal midline across overlapping regions of the Central Nervous System (CNS). Wnt-3a signaling is essential for various morphogenetic events, including embryonic patterning, cell determination, cell proliferation, CNS development, and cytoskeletal formation. Like other members of this family, Wnt-3a contains a highly conserved lipid-modified, cysteine-rich domain that is essential for cell signaling. During a biochemical process called the canonical Wnt pathway, Wnt family members bind to and activate, seven-pass transmembrane receptors of the Frizzled family, ultimately leading to the disruption of β-catenin degradation. Intracellular accumulation of β-catenin increases translocation of the protein into the nucleus, where it binds to TCF/LEF transcription factors to promote gene expression. Lack of Wnt signaling disrupts transcriptional activation of tumor suppressor genes, and has been shown to result in neoplastic transformation, oncogenesis, and human degenerative diseases.
Wnt-9b
Formerly known as Wnt-15 or Wnt-14b, Wnt-9b is a secreted glycoprotein belonging to the Wnt family of signaling proteins that are critically involved in maintaining the integrity of both embryonic and adult tissues. Wnt-9b is primarily expressed in adult kidneys and during late embryogenesis, and shares with other Wnt family members the same highly conserved lipid-modified, cysteine-rich domain essential for cell signaling. As is true for most Wnt family members, Wnt-9b functions through the biochemical process known as the canonical Wnt pathway. In this process, Wnt proteins bind to and activate seven-pass transmembrane receptors of the Frizzled family, and ultimately result in the disruption of β-catenin degradation. Intracellular accumulation of β-catenin increases translocation of the protein into the nucleus, where it binds to TCF/LEF transcription factors to promote the expression of numerous genes. In this manner, Wnt signaling induces and maintains transformed phenotype, and, in certain embryonic cell lines, supports self-renewal in the absence of significant differentiation. While increased Wnt/β-catenin signaling is associated with tumorigenesis in a diverse set of human cancers, lack of Wnt signaling disrupts transcriptional activation of tumor suppressor genes, and has been shown to result in neoplastic transformation, oncogenesis, and human degenerative diseases. Altered Wnt-9b expression has been shown to result in the underdevelopment of the kidneys, and incomplete lip and cleft fusion in mice.